Treadmill exercise ameliorates motor dysfunction through inhibition of Purkinje cell loss in cerebellum of valproic acid-induced autistic rats
Article information
Abstract
Autism is a complex developmental disorder with impairments in social interaction, communication, repetitive behavior and motor skills. Exercise enhances cognitive function, ameliorates motor dysfunction, and provides protective profits against neurodegeneration. In the present study, we evaluated the effect of treadmill exercise on the motor coordination and Purkinje cell loss in relation with reactive astrocytes and microglial activation in the cerebellum using valproic acid (VPA)-induced autism rat model. On the 12th day of pregnancy, the pregnant rats in the VPA-exposed group received intraperitoneal injections of 600-mg/kg VPA. After birth, the rat pups were divided into four groups: the control group, the exercise group, the VPA-treated group, the VPA-treated and exercise group. The rat pups in the exercise groups were forced to run on a treadmill for 30 min once a day, 5 times a week for 4 weeks. In the present results, motor balance and coordination was disturbed by induction of autism, in contrast, treadmill exercise alleviated motor dysfunction in the autistic rats. Purkinje cell loss, reactive astrocytes, and microglial activation were occurred by induction of autism, in contrast, treadmill exercise enhanced survival rate of Purkinje neurons through inhibition of reactive astrocytes and microglia in the autistic rats. The present study showed that exercise may provide a potential therapeutic strategy for the alleviation of motor dysfunction in autistic patients.
INTRODUCTION
Autism is a complex developmental disorder with impairments in social interaction, communication, repetitive behavior and motor skills. Clinical studies of children exposed to valproic acid (VPA) in utero have characterized a fetal valproate syndrome with symptoms similar to autism (Williams et al., 2001). Also, prenatal exposure to VPA in rodents lead to neurodevelopmental deficits that are similar to the motor and cognitive deficits seen in human diagnosed with autism (Wagner et al., 2006).
Several studies reported cerebellar abnormality in autistic samples (Bauman and Kemper, 2005; Courchesne et al., 1994). Purkinje neurons in the cerebellum are associated with movement, coordination, and cognitive function (Seo et al., 2010). Ninety-five percent of autistic cases examined at autopsy showed cerebellar pathology including a reduction of Purkinje neurons (Bailey et al., 1998; Kemper and Bauman, 2002).
Neuroglial cells such as astrocytes and microglia play important roles in neuronal function and homeostasis (Aloisi, 2001; Dong and Benveniste, 2001). Reactive astrocyte is very sensitive markers of neuronal damage in the brain (Devinsky et al., 2013). Similarly, microglial activation is an important factor in the neuroglial responses to injury or dysfunction (Streit and Xue, 2009). Immunocytochemical studies of brain tissues from autistic patients showed marked activation of microglia and astroglia as compared with controls (Li et al., 2009). The magnitude of astrogliosis can be assessed by using immunocytochemistry for glial fibrillary acidic protein (GFAP). CD11b is the surface marker located on the plasma membrane of microglia, the expression of which is increased during microglial activation (Singh et al., 2011).
Exercise enhances cognitive function, ameliorates motor dysfunction, and provides protective profits against neurodegeneration (Heo et al., 2014; Lee et al., 2014; Seo et al., 2010). Exercise may attenuate neurological impairments in various neuropsychiatric disorders, including autism (Kim et al., 2013; Petrus et al., 2008). However, the effect of exercise on the survival of Purkinje neurons in relation with reactive astrocytes in autism has not been clarified. In the present study, we evaluated the effect of treadmill exercise on the motor coordination and Purkinje cell loss in relation with reactive astrocytes and microglial activation in the cerebellum using VPA-induced autism rat model.
MATERIALS AND METHODS
Animals and treatments
This study was performed in accordance with the guidelines of the National Institutes of Health and the Korean Academy of Medical Sciences. Female Sprague-Dawley rats (180±10 g, 8 weeks old, n=8) were allowed to mate with male rats for 24 hr. The female rats were then individually housed in plastic home cages under the controlled temperature (20°C±2°C) and a light–dark cycle consisting of 12 hr of light and 12 hr of darkness (lights on from 7:00 a.m. to 7:00 p.m.). Food and water were made available ad libitum.
The pregnant rats were divided into two groups: the control group and the VPA-exposed group (n=4 in each group). On the 12th day of pregnancy, the pregnant rats in the VPA-exposed group received intraperitoneal injections of 600-mg/kg VPA (Sigma Chemical Co., St. Lois, MO, USA) dissolved in 1-mL saline, and the pregnant rats in the control group were treated with saline in the same volume and on the same schedule.
After birth, the rat pups were divided into four groups: control group, exercise group, VPA-treated group, VPA-treated and exercise group (n=10 in each group).
Treadmill exercise protocol
The rat pups in the exercise groups were forced to run on a treadmill for 30 min once a day, 5 times a week for 4 weeks, starting postnatal day 42. Exercise load for the exercise groups consisted of running at a speed of 3 m/min for the first 5 min, at a speed of 5 m/min for the next 5 min, and then at a speed of 8 m/min for the last 20 min, with the 0° inclination.
Rota-rod test
We performed a rota-rod (Harvard Apparatus, Holliston, MA, USA) test to measure motor balance and coordination at 4 weeks after treadmill exercise. Each rat was placed in a separate compartment on the rotating rod (diameter, 7 cm). The velocity of the rod was set at a constant 5 rpm. Latency until fall was automatically recorded by magnetic trip plates. To eliminate stress and fatigue, rats were given a maximum cutoff latency of 180 sec.
Tissue preparation
The animals were sacrificed immediately after determining the rota-rod test. To prepare the brain slices, the animals were fully anesthetized with diethyl ether, transcardially perfused with 50-mM phosphate-buffered saline (PBS), and fixed with a freshly prepared solution consisting of 4% paraformaldehyde in 100-mM phosphate buffer (pH, 7.4). Brains were dissected, and storage overnight same fixative, then it was transferred to 30% sucrose for cryoprotection. Sagittal sections of 40-μm thickness in each section of cerebellum were made with a freezing microtome (Lieca, Nussloch, Germany).
Immunohistochemistry
In order to detect Purkinje neurons and reactive astrocytes in the cerebellar vermis, immunohistochemistry for calbindin and GFAP was performed, according to the previously described method (Lee et al., 2014). Free-floating tissue sections were first incubated in 3% H2O2 for 30 min to block the endogenous peroxidase activity. The sections were incubated in blocking solution (1% bovine serum albumin and 10% horse serum in 0.05-M PBS) for 2 hr at room temperature, and were then incubated overnight with mouse anticalbindin D-28k antibody (1:2,000; Sigma Chemical Co.) or with mouse monoclonal anti-GFAP antibody (1:3,000; Chemicon, Temecula, CA, USA) at 4°C. The next day, the sections were incubated for 1 hr with anti-mouse secondary antibody (1:200; Vector Laboratories, Burlingame, CA, USA) at room temperature, and were subsequently incubated with a Vector Elite ABC kit (Vector Laboratories) for 1 hr at room temperature. Between all steps, the sections were thoroughly washed 3 times in 0.05-M PBS. Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.05% 3,3′-diaminobenzidine and 0.01% H2O2 in 0.05-M Tris buffer (pH, 7.6) for approximately 5 min. The visualized sections were mounted on gelatin-coated slides and coverslipped with mounting medium.
Western blotting
Western blot for the GFAP and CD11b was performed, according to the previously described method (Kim et al., 2013). The cerebellum tissues were dissected and collected, and then were immediately frozen at −70°C. The right hemisphere were homogenized on ice, and lysed in a lysis buffer containing 50-mM N-2-hydroxyethylpiperazine-N-2-ethanesulfonic acid (pH 7.5), 150-mM NaCl, 10% glycerol, 1% Triton X-100, 1-mM phenylmethylsulfonyl fluoride, 1 mM ethyleneglycol-bis-(b-aminoethylether)-N,N,N′,N′-tetraacetic acid, 1.5-mM MgCl2-6H2O, 1-mM sodium orthovanadate, and 100-mM sodium fluoride. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Hercules, CA, USA). Protein samples (30 μg) were separated on sodium dodecyl sulfate-polyacrylamide gel and transferred onto a nitrocellulose membrane. The membranes were incubated with 5% skim milk in Tris-buffered saline containing 0.1% Tween-20 and then incubated overnight at 4°C with the following primary antibodies: mouse anti-β-actin antibody, anti-GFAP and anti-CD11b (1:1,000; Santa Cruz Biotechnology, Santa Cruz, CA, USA). Subsequently, membranes were incubated for 1 hr with attempt secondary antibodies (1:2,000; Vector Laboratories), and ban detection was performed using the enhanced chemiluminescence detection kit (Santa Cruz Biotechnology).
Data analysis
Cell counting and optical density measurements for calbindin and GFAP expressions were performed using Image-Pro Plus software (Media Cybernetics, Silver Spring, MD, USA) attached to a light microscope (Olympus, Tokyo, Japan). The number of Purkinje-positive cells was quantified in a field with dimensions of 1,000 μm×400 μm in the regions of the cerebellar vermis. The density of GFAP-positive reactive astrocytes was assessed in a quantitative fashion, according to a microdensitometrical method based on optical density. To compare the relative expression of GFAP and CD11b, the detected bands were calculated densitometrically using Molecular Analyst, version 1.4.1 (Bio-Rad).
Statistical analysis was performed using one-way analysis of variance followed by Duncan post hoc test, and the results are expressed as the mean±standard error of the mean. Significance was set as P<0.05.
RESULTS
Effect of treadmill exercise on the motor function in the rota-rod test
The latencies to fall of the rota-rod test are presented in Fig. 1. The results showed that motor coordination and balance was disturbed by VPA-induced autism (P<0.05), whereas treadmill exercise ameliorated motor dysfunction in the VPA-induced autistic rats (P<0.05).
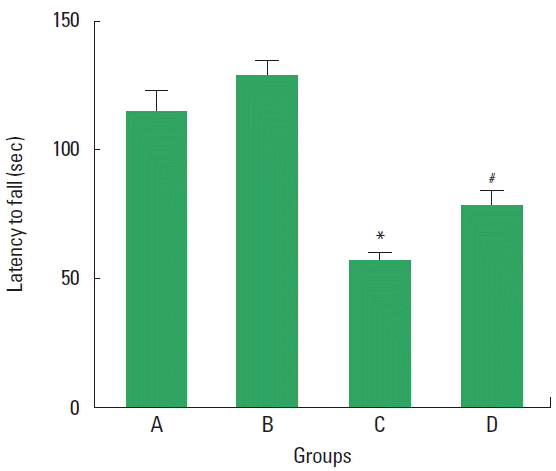
Effect of treadmill exercise on the motor function in the rota-rod test. A, control group; B, exercise group; C, valproic acid (VPA)-treated group; D, VPA-treated and exercise group. Values are presented as mean±standard error of the mean. *P<0.05 compared to the control group. #P<0.05 compared to the VPA-treated group.
Effect of treadmill exercise on Purkinje neurons in the cerebellar vermis
Photomicrographs of calbindin-positive Purkinje neurons in the anterior region of cerebellar vermis are presented in Fig. 2. These results showed that Purkinje neuronal loss in the cerebellar vermis was increased by VPA-induced autism (P<0.05), whereas treadmill exercise inhibited autism-induced Purkinje cell loss.

Effect of treadmill exercise on the number of Purkinje neurons in the cerebellal vermis. Upper panel: Photomicrographs of calbindin-positive cells in the anterior region of cerebellar vermis (immunohistochemistry). The scale bar represents 50 μm. Lower panel: The number of calbindin-positive cells in the anterior region of cerebral vermis. A, control group; B, exercise group; C, valproic acid (VPA)-treated group; D, VPA-treated and exercise group. Values are presented as mean±standard error of the mean. *P<0.05 compared to the control group. #P<0.05 compared to the VPA-treated group.
Effect of treadmill exercise on reactive astrocytes in the cerebellar vermis
Photomicrographs of GFAP-positive cells in the anterior region of cerebellar vermis are presented in Fig. 3. Expression of reactive astrocytes in the cerebellar vermis was enhanced by VPA-induced autism (P<0.05), whereas treadmill exercise suppressed autism-induced expression of reactive astrocytes (P<0.05).

Effect of treadmill exercise on the expression of reactive astrocytes in the cerebellar vermis. Upper panel: Photomicrographs of glial fibrillary acidic protein (GFAP)-positive cells in the anterior region of cerebellar vermis (immunohistochemistry). The scale bar represents 50 μm. Lower panel: The optical density of GFAP-positive cells in the anterior region of cerebellar vermis. A, control group; B, exercise group; C, valproic acid (VPA)-treated group; D, VPA-treated and exercise group. Values are presented as mean±standard error of the mean. *P<0.05 compared to the control group. #P<0.05 compared to the VPA-treated group.
Effect of treadmill exercise on reactive astrocytes in the cerebellum
GFAP expression in the cerebellum is presented in Fig. 4. Expression of reactive astrocytes in the cerebellum was enhanced by VPA-induced autism (P<0.05), whereas treadmill exercise suppressed autism-induced expression of reactive astrocytes (P<0.05).
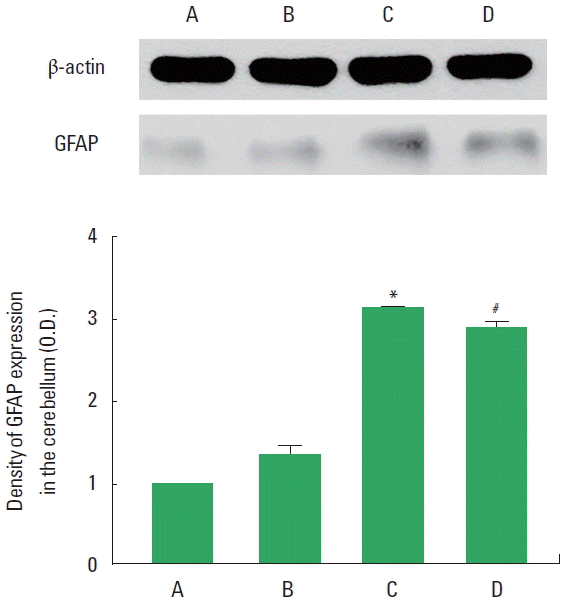
Effect of treadmill exercise on the glial fibrillary acidic protein (GFAP) expression in the cerebellum. Upper panel: Representative expression of GFAP and β-actin in the cerebellum. Lower panel: Relative GFAP expression in each group. A, control group; B, exercise group; C, valproic acid (VPA)-treated group; D, VPA-treated and exercise group. Values are presented as mean±standard error of the mean. *P<0.05 compared to the control group. #P<0.05 compared to the VPA-treated group.
Effect of treadmill exercise on microglial activation in the cerebellum
CD11b expression in the cerebellum is presented in Fig. 5. Microglial activation in the cerebellum was increased by VPA-induced autism (P<0.05), whereas treadmill exercise inhibited autism-induced microglial activation (P<0.05).
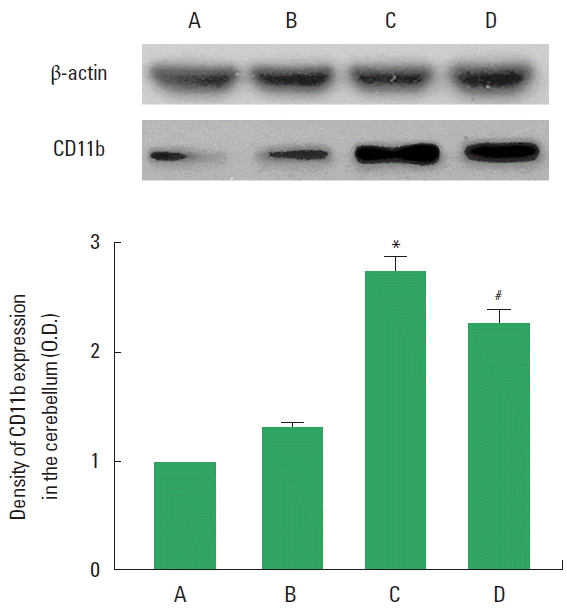
Effect of treadmill exercise on the CD11b expression in the cerebellum. Upper panel: Representative expression of CD11b and β-actin in the cerebellum. Lower panel: Relative CD11b expression in each group. A, control group; B, exercise group; C, valproic acid (VPA)-treated group; D, VPA-treated and exercise group. Values are presented as mean±standard error of the mean. *P<0.05 compared to the control group. #P<0.05 compared to the VPA-treated group.
DISCUSSION
Prenatal exposure to chemicals such as alcohol, lead, and VPA, and radiation are well known to induce developmental abnormalities in the central nervous system of children (Carpenter et al., 2001; Smalley et al., 2002). Anticonvulsant medication during pregnancy increased the risk of autism and sodium valproate is the most commonly associated drug with autistic disorder (Rasalam et al., 2005). Several studies demonstrated administration of VPA to pregnant rats showed autism-like symptoms, including neuron loss in the brainstem nuclei and cerebellum (Ingram et al., 2000) and behavioral alterations (Stanton et al., 2007).
In the present study, the pregnant rats in the VPA-exposed group received injections of 600-mg/kg VPA on the 12th day of pregnancy, according to previously described method (Stanton et al., 2007). This study demonstrated that a dose of 600-mg/kg VPA induced loss of cerebellar Purkinje cells and impairment of motor coordination, which demonstrated that this dosage was sufficient to induced autism in the rat pups.
The cerebellum is important brain area in motor coordination and balance and it has been recognized as a prominent contributor to a wide array of cognitive and emotional functions (Tiemeier et al., 2010). Disturbance of Motor function is important aspect of the description of autism (Vernazza-Martin et al., 2005).
We used the rota-rod test for the evaluation of motor abnormality and imbalance. In the present study, the latency to fall in the rota-rod test was decreased in the VPA-induced autistic rats. However, treadmill exercise increased the latency in the autistic rats. The present results demonstrated that motor balance and coordination was disturbed by induction of autism, in contrast, treadmill exercise alleviated motor dysfunction in the autistic rats.
Neuroglial cells, such as astrocytes and microglia, play modulate neuronal function and homeostasis (Aloisi, 2001). However, during reactive astrocytes and microglial activation secondary to injury or in response to neuronal dysfunction, several factors that evoke inflammatory responses are secreted (Rosenberg, 2002). Therefore, reactive astrocytes and microglia activation induce neuronal and synaptic changes that are likely to contribute to central nervous system dysfunction during disease processes. In the brains of autistic patients, enhanced astrocyte reactivity with neuronal loss was observed in the cerebellum (Streit and Xue, 2009). Seo et al. (2010) reported that treadmill exercise enhanced the survival rate of Purkinje cells in the cerebellar vermis following traumatic brain injury.
In the present study, the number of calbindin-stained Purkinje neurons was decreased and the expressions of GFAP and CD11b were increased in the cerebellum of autistic rats. However, treadmill exercise increased the number of calbindin-stained Purkinje neurons and suppressed the expressions of GFAP and CD11b in the autistic rats. The present results demonstrated that Purkinje cell loss, reactive astrocytes, and microglial activation were occurred by induction of autism, in contrast, treadmill exercise enhanced survival rate of Purkinje neurons through inhibition of reactive astrocytes and microglia in the autistic rats.
We demonstrated that VPA-induced autism increased Purkinje cell loss with activation of reactive astrocytes and microglia in the cerebellum, resulting in decreased motor coordination. However treadmill exercise suppressed Purkinje cell loss via down-regulation of reactive astrocytes and microglia, and then ameliorated motor dysfunction in autistic rats. The present study showed that exercise may provide a potential therapeutic strategy for the alleviation of motor dysfunction in autistic patients.
ACKNOWLEDGMENTS
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (NRF-2013 S1A5B5A07048045).
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.