INTRODUCTION
Schizophrenia is a serious psychiatric disorder with 0.5 to 1.0 percent prevalence worldwide (Ross et al., 2006). Schizophrenia causes particular collapse in personality, such as thought without any clouding of consciousness, affection, perception, volition, and motor behavior, etc. The exact mechanisms of schizophrenia have not been identified, however previous studies have suggested that neurochemical dysfunction is important clue for schizophrenia, such as dopamine dysfunction, dopamine-glutamate imbalance, and glutamate receptor hypofunction (Carlsson et al., 1990, 1999; Deutsch et al., 2002). According to the Coyle (2006), there is new evidence to suspect that the abnormality in NMDA receptor function may contribute to schizophrenia’s symptoms that are resistant to antipsychotic medication.
Glutamate is the primary excitatory neurotransmitter in the mammalian brain. Glutamate acts postsynaptically on the three families of ionotropic receptors, such as N-methyl D-aspartate (NMDA) receptor, a-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA) receptor, and kainate receptor (McBain and Mayer, 1994). Excitatory action of glutamate is prominent particularly in the hippocampus, out layer of the cerebral cortex, and substantia gelationsa of the spinal cord. Excitatory action of glutamate is implicated in many physiological processes including learning and memory (Hudspith. 1997).
Dopamine is synthesized first by hydroxylation of the amino acid L-tyrosine to 3,4-dihydroxy-L-phenylalanine by the enzyme tyrosine hydroxylase (TH), and then it is decarboxylated by L-amino acid decaboxylase to dopamine (Dunkley et al., 2004).
Brain-derived neurotrophic factor (BDNF) is one of the neurotrophin families which interact with high affinity protein tyrosine kinase receptor (Trk). BDNF plays key roles in early embryogenensis by enhancing the differentiation of neuronal precursors as well as being a critical component of early neural tube development in the chick (Kernie et al., 2000). BDNF expression is altered in various brain diseases, and BDNF is also involved in eating behavior and physical activity (Trpia-Arancibia et al., 2004).
Schizophrenia patients show deficit in a variety of cognitive function, and this deficit is associated with dysfunction of network in brain areas including the frontal and temporal cortex, the hippocampus, and the subcortial regions (Kuperberg and Heckers, 2000). In particularly, dysfunction of prefrontal cortex is associated with working memory deficit (Manoach. 2003), and this abnormality is predominant in schizophrenia (Meyer-Lindenberg et al., 2002).
Administration of NMDA receptor antagonists and dopaminergic agonists to animals is the most widely used animal models for schizophrenia (Rung et al., 2005). MK-801, also known as dizocilpine maleate, is a non-competitive NMDA receptor antagonist, and this agent has been used for inducing schizophrenia-like symptoms in animals.
Exercise is known to protect neurons from various brain insults, activates neuronal cells, promotes neurogenesis, increases brain plasticity, and enhances cognitive function (Collins et al., 2009; van Praag, 2009; Weuve et al., 2004). Many studies have revealed that exercise improves symptoms of neuropsychiatric disorders (Cho et al., 2013; Jee et al., 2008; Seo et al., 2013), however the effect of treadmill exercise on the schizophrenia in relation with NMDA receptor expression have not been evaluated. In the present studies, the effect treadmill running on the NMDA receptor expression was investigated using MK-801-induced schizophrenia mice.
MATERIALS AND METHODS
Materials
MK-801 was dissolved in saline (0.9% NaCl solution) as master stock solution (1 mg/mL). The stock solution was diluted with saline to the final concentrations (0.6 mg/kg) for systemic administration by intraperitoneal injection. Dilution was designed to give a final injection volume of 100 μL per 10 grams of animal weight for each injection of MK-801.
Experimental animals and design
Male C57BL/6 mice weighing 27±3 g (6 weeks of age) were used for this experiment. The animals were housed under standard housing conditions with plastic cages. The animals were housed under controlled temperature (23±2°C) and lighting (08:00 to 20:00) conditions, and were supplied with food and water ad libitum. The experimental procedures were performed in accordance with the animal care guidelines of the National Institutes of Health (NIH) and the Korean Academy of Medical Sciences. The mice were randomly divided into three groups (n=10 in each group): control group, MK-801 injection group, MK-801 injection and treadmill exercise group.
Exercise protocol
The mice in the treadmill exercise group were made to run on a motor-driven treadmill for 1 hour once a day during 2 weeks. The exercise load consisted of running at the speed of 2 meters/min for the first 5 min, 5 meters/min for the next 5 min, and then 8 meters/min for the last 50 min, with the 0° inclination. The mice in the control group and in the MK-801 injection group were left in the treadmill, without running, for the same duration as the exercise group.
Tissue preparation
The mice were sacrificed immediately after last treadmill exercise. The animals were anesthetized using Zoletil 50® (10 mg/kg, i.p.; Vibac Laboratories, Carros, France), transcardially perfused with 50 mM phophate-buffered saline (PBS), and fixed with a freshly prepared solution consisting of 4% paraformaldehyde in 100 mM phophate buffer (PB, pH 7.4). Brain was dissected, post-fixed in the same fixative overnight and tranferred to 30% sucrose for cryoprotection. 40 μm thick coronal sections were made using a freezing microtome (Leica, Nussloch, Germany). Ten slice sections on average were collected from each mouse.
NMDA receptors immunohistochemistry
To visualize NMDA receptor expression, immunohistochemistry for NMDA receptor was performed, as the previously described method (Eleore et al., 2005). The sections were selected from each brain and were incubated overnight with anti-NMDR1 primary antibody (1:1,000; Vector Laboratories, Burlingame, CA, USA), and then with biotinylated second antibody for 1 h. The second antibody was amplified with the Vector Elite ABD kit® (1:100; Vector Laboratories). Anti-biotin-avidin-peroxidase complexes was visualized using 0.03% 3,3-diaminobenzidine (DAB), and the section was mounted onto gleatin-coated slides. The slides were air-dried overnight at room temperature, and coverslips were mounted using Permount® (Fisher Scientific, Fair Lawn, NJ, USA).
Tyrosine hydroxylase immunohistochemistry
For immunolabeling of TH in the striatum, TH immunohistochemistry was performed as the previously described method (Cho et al., 2013; Kim et al., 2005). Free-floating tissue sections were incubated overnight with mouse anti-TH antibody (1:1,000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) and the sections were then incubated for 1 h with biotinylated anti-mouse secondary antibody (1:2,000, Vector Laboratories). The sections were subsequently incubated with avidin-biotin-peroxidase complex (Vector Laboratories) for 1 h at room temperature. Immunoreactivity was visualized by incubating the sections in a solution consisting of 0.05% DAB and 0.01% H2O2 in 50 mM Tris-buffer (pH 7.6) for approximately 3 min. The sections were then washed three times with PBS and mounted onto gelatine-coated slides. The slides were air-dried overnight at room temperature, and coverslips were mounted using Permount® (Fisher Scientific).
Western blot for BDNF expression
BDNF expression was determined by western blot as the previously described method (Kim et al., 2010). The hippocampal tissues were collected, and then are immediately frozen at −70°C. The hippocampal tissues was homogenized on ice, and lysed in a lysis buffer containing 50 mM Tris-HCl (pH 7.5), 150 mM NaCl, 0.5% deoxycholic acid, 1% Nonidet P40, 0.1% SDS, 1 mM PMSF, and 100 mg/mL leupeptin. Protein content was measured using a Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein of 30 μg was separated on SDS-polyacrylamide gels and transferred onto a nitrocellulose membrane. Mouse β actin antibody (1:3,000; Santa Cruz Biotechnology), rabbit BDNF antibody (1:1,000; Santa Cruz Biotechnology).
Horseradish peroxidase-conjugated anti-rabbit antibody for BDNF was used as the secondary antibody. Experiment was performed in normal lab conditions and at room temperature except for transferred membrane. Membrane transfer was performed at 4°C with the cold pack and prechilled buffer. Band detection was performed using the enhanced chemiluminescence (ECL) detection kit (Santa Cruz Biotechnology). To compare relative expression of proteins, detected bands were calculated densitometrically using Molecular Analyst™ version1.4.1 (Bio-Rad).
Data analysis
For confirming the expression of BDNF, the detected bands were calculated densitometrically using Molecular Analyst™, version 1.4.1. The number of NMDA-positive cells were counted hemi-laterally under a light microscope (Olympus, Tokyo, Japan), and they were expressed as the number of cells per square millimeter. The area of the dentate gyrus was measured by Image-Pro® Plus image analysis system (Media Cyberbetics Inc., Silver Sprng, MD, USA). To estimate the TH-staining density, the optical densities were corrected for nonspecific background density, which was measured in completely denervated parts of the striatum. The ratio of TH-positive fiber density in the striatum was calculated as follows: optical density in the lesion side/optical density in the intact side.
All data were analyzed using the statistical software SPSS (version 15.0). The data was analyzed with one-way ANOVA and then Duncan’s post-hoc tests. All values are expressed as the mean±standard error of the mean (SEM). The P value<0.05 was considered significant.
RESULTS
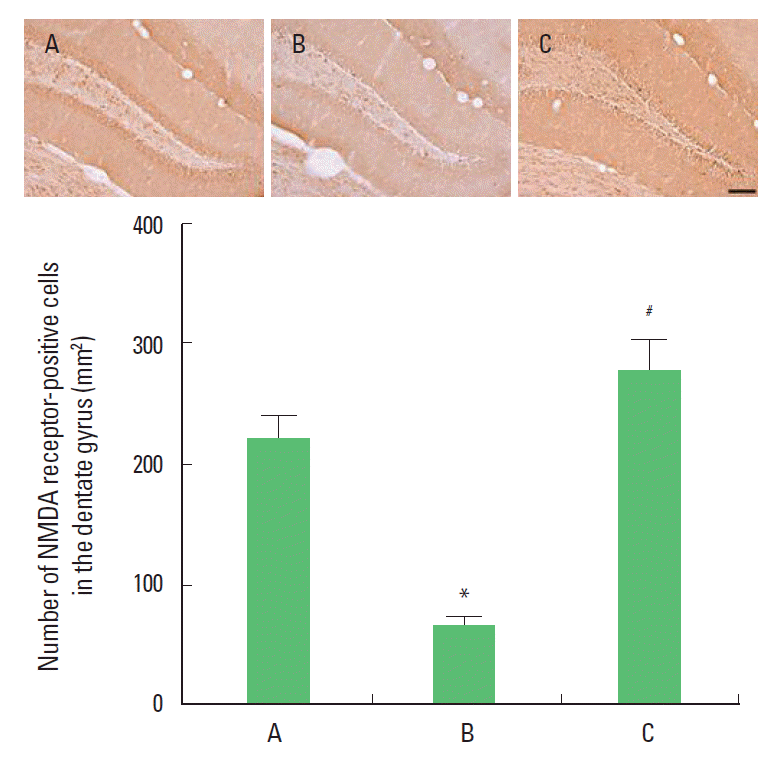
Effect of treadmill exercise on the number of NMDA receptor-positive cells in the dentate gyrus
Photomicrographs of the NMDA receptor-positive cells in the dentate gyrus are presented Fig. 1. The number of NMDA receptors-positive cells was 220.40±18.91/section in the control group, 64.01±7.93/section in the MK-801 injection group, and 275.92±24.74/section in the MK-801 injection and treadmill exercise group. The present results showed that the number of NMDA receptor-positive cells in the dentate gyrus of the MK-801 injection group was decreased compared to the control group (P<0.05). In contrast, the number of NMDA receptor-positive cells in the dentate gyrus of the treadmill exercise group was increased (P<0.05).
Effect of treadmill exercise on the number of NMDA receptor-positive cells in the prefrontal cortex
Photomicrographs of the NMDA receptor-positive cells in the prefrontal cortex are presented Fig. 2. The number of NMDA receptors-positive cells were 1,724.10±47.50/section in the control group, 1,022.50±28.03/section in the MK-801 injection group, 1,355.43±22.48/section in the MK-801 injection and treadmill exercise group. The present results showed that the number of NMDA receptor-positive cells in the prefrontal cortex of the MK-801 injection group was decreased compared to the control group (P<0.05). In contrast, the number of NMDA receptor-positive cells in the prefrontal cortex of the treadmill exercise group was increased (P<0.05).
Effect of treadmill exercise on the TH expression in the striatum
Photomicrographs of the TH in the striatum are presented Fig. 3. The density of TH was 87.66±3.22 in the control group, 104.41±3.24 in the MK-801 injection group, 89.90±0.79 in the MK-801 injection and treadmill exercise group. The present result showed that the TH expression in the striatum of the MK-801 injection group was increased compared to the control group (P<0.05). In contrast, TH expression in the striatum of the treadmill exercise group was increased (P<0.05).
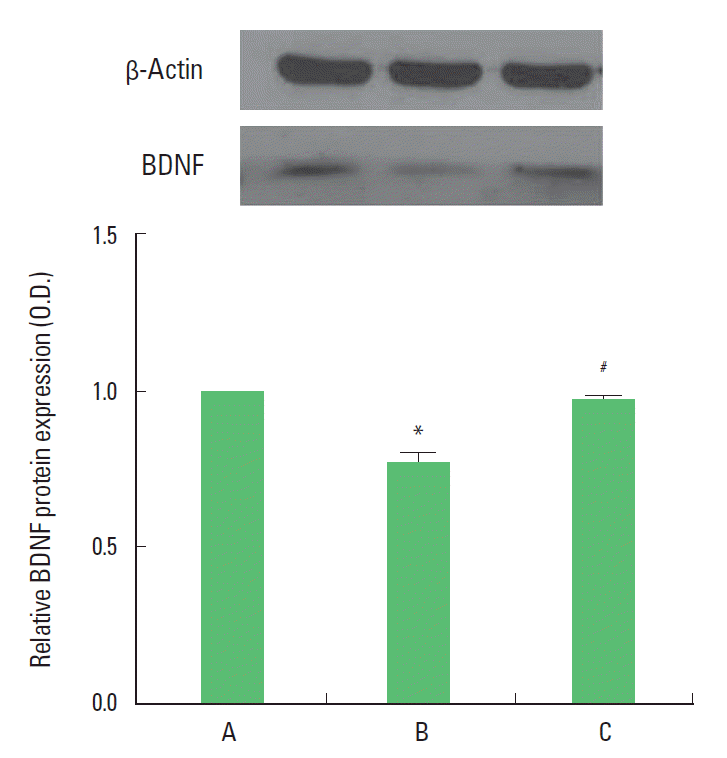
Effects of treadmill exercise on the BDNF expression in the hippocampus
The BDNF (14 kDa) protein expressions in the hippocampus are presented Fig. 4. When the level of BDNF in the control group was set at 1.00, the level of BDNF was 0.77±0.03 in the MK-801 injection group, 0.97±0.01 in the MK-801 injection and treadmill exercise group. The present result showed that the expression of BDNF in the hippocampus of the MK-801 injection group was decreased in compared to the control group (P<0.05). In contrast, BDNF expression in the hippocampus of the treadmill exercise group was increased (P<0.05).
DISCUSSION
Many studies have indicated that NMDA receptor hypofunction may contribute to the pathophysiology of schizophrenia and induces cognitive and behavior abnormalities (Duncan et al., 2004; Kristiansen et al., 2007; Newcomer and Krystal, 2001). NMDA receptor causes diverse functions in the sensory and motor system (Daw et al., 1993). Duncan et al. (2006) reported that NMDA receptor hypofunction is related to the increased locomotor activity. NMDA receptor antagonists induce hyperactivity that resembles schizophrenia-like symptom (Leriche et al., 2003). Mice with reduced NMDA receptor expression display behavioral abnormalities, including deficit in social interaction (Mohn et al., 1999). Postmortem study reported that NMDA receptor subunit was decreased in the brain regions of schizophrenia patients, especially in the hippocampus and prefrontal cortex (Harrison et al., 2003).
In the present results, NMDA receptor expression was suppressed in the prefrontal cortex and hippocampus of the MK-801-iduced schizophrenia mice. The present results also showed that treadmill exercise increased NMDA receptor expression in the hippocampus and prefrontal cortex.
Previous studies revealed that exercise lead to change in NMDA receptor expression. Kitamura et al. (2003) reported that wheel exercise may activate NMDA receptors in the hippocampus, and exercise increased the phosphorylated form of NMDA receptor (Dietrich et al., 2005). NMDA receptor exerts an important role in neuronal plasticity and synaptic signaling through a fine control of its channel activity (Dietrich et al., 2005). Exercise also activates NMDA receptor in the hippocampus (Dietrich et al., 2005; Vaynman et al., 2003).
Zafra et al. (1990) suggested that the synthesis of neurotrophic factors such as BDNF and nerve growth factor (NGF) in the brain is regulated by neuronal activity via NMDA receptor. BDNF expression was decreased in the several neuropsychiatric disorders, including schizophrenia and depression (Knable et al., 2004). NMDA receptor antagonist MK-801 reduced expression of BDNF in the hippocampus (Fumagalli et al., 2003).
In the present study, BDNF expression was decreased hippocampus of the MK-801-iduced schizophrenia mice. The present results also showed that treadmill exercise increased BDNF expression in the hippocampus.
BDNF-improving effect of physical exercise has been reported in many other studies (Garza et al., 2004; Griebach et al., 2004; Huang et al., 2006). Exercise-induced enhancement of BDNF expression in the hippocampus inhibited age-dependent deterioration of short-term and spatial working memories (Kim et al., 2010). High level of BDNF also alleviates spatial working memory impairment in attention deficit/hyperactivity disorder (ADHD) rats (Kim et al., 2011).
NMDA receptor antagonists, such as MK-801 or phencyclidine induced behavioral abnormalities which are accompanied by an increment in dopaminergic and serotonergic neuronal activity in various brain regions (Löscher et al., 1991). The glutamatergic system and dopaminergic system are tightly linked to each other. Reduced glutamate function, induced by MK-801, may cause some elevation of dopamine release (Carlsson et al., 2000). Larulle et al. (2003) reported that deficits in glutamate transmission might lead to the increment of dopamminergic biological marker associated with schizophrenia. TH is the first and rate-limiting enzyme in catecholamine synthesis and it catalyses the hydroxylation of l-tyrosine to DOPA (Dunkley et al., 2004). TH activity is known to be gradually decreased following the loss of dopamine neurons (Yoon et al., 2007), as a result, TH represents the rate of dopamine synthesis (Howes et al., 2009).
The present results showed that the density of TH in the striatum was increased by MK-801 injection. Dopamine hyperactivity in the striatum is associated with the prodromal signs of schizophrenia (Howes et al., 2009). In the present results, treadmill exercise suppressed overactivity of density of TH. Lindgren et al. (2000) observed that NMDA receptor activity decreased dopamine synthesis in the stiatum. As mentioned above, it is possible that the suppression of TH is due to the increment of NMDA receptor by exercise.
The present study showed that down-regulation of NMDA receptor demonstrated schizophrenia-like parameters, meanwhile treadmill running improved schizophrenia-related parameters through enhancing NMDA receptor expression. Based on the present results, it can be suggested that treadmill exercise may be used as the one of the important strategies for the prevention and treatment of schizophrenia patients.