INTRODUCTION
Routine exercise training of moderate intensity is known to enhance immune function (Walsh et al., 2011) and reduce risk of mortality (Liu et al., 2012; Sabia et al., 2012) cardiovascular diseases, obesity, and type 2 diabetes (Pedersen, 2006). However, an acute bout of prolonged (>1.5 hr) and/or strenuous exercise (55%–75% maximal O2 uptake) has also been shown to provoke muscle damage/soreness (Clarkson and Hubal, 2002), and elicit an acute inflammatory (Nieman et al., 2012), oxidative stress (Fisher et al., 2011) during the postexercise recovery period. Exhaustive and/or unaccustomed exercises (particularly those involving high intensity muscle contractions) are known to induce temporary muscle damage. This damage includes a series of events occurring. It causes damage to the sarcolemma and the muscle cell membrane. This damage results in the release of biochemical markers of muscle damage including (e.g., lactate dehydrogenase, creatine kinase, and myoglobin) (Torres et al., 2012). Exercise-induced muscle damage is classified into primary and the secondary damage (Howatson and van Someren, 2008). Primary muscle damage is related to morphological changes, including sarcomere (Z-disc, I, and A band), sarcolemma, sarcoplasmic reticulum, and cytoskeletal elements (Clarkson and Hubal, 2002). Secondary muscle damage occurs due to impaired calcium homeostasis and the inflammatory response (Tidball, 2005).
According to recent studies, contracting muscles release various cytokines called myokines. This finding has opened a new insight on the role of cytokines in modulating the muscle’s metabolic and immunological responses to exercise. Different observations in cytokine responses to exercise are predominantly related to the intensity, mode and duration of exercise in addition to the participants’ fitness levels, although differences in sensitivity and specificity of the assays should be considered. Acute bouts of intense or prolonged resistance exercise can induce an inflammatory response (Colbert et al., 2004). IL-6 could have a role during the inflammatory process in the enhancement of muscle damage or during the healing process. Today it is well established that IL-6 is secreted by contracting skeletal muscle in the absence of muscle damage and that it plays important roles in the metabolic regulation (Steinbacher and Eckl, 2015).
CRP is an acute phase protein which increases remarkably during infection, inflammation and tissue damage (Borish and Steinke, 2003). It is mainly produced and secreted into circulation by liver in response to circulatory inflammatory mediators (Shoelson and Lee, 2006). CRP level is the best indicator to determine tissue damage because CRP increased immediately at early phase of tissue deterioration (Simopoulos, 2006). Recent studies reveal that CRP not only is an inflammatory mediator, but it plays very important role in inflammation (Peake et al., 2005).
Pentraxins are a group of multimeric proteins that are involved in acute immunologic responses. Short pentraxins (CRP and amyloid P protein) are synthesized by the liver in response to inflammatory signals. PTX-3 is the prototype of long pentraxins (molecular weight, 440 kDa) and is produced in response to inflammatory signals in various organs and tissues including vascular endothelial cells, smooth muscle cells, macrophages, adipose tissue, and liver (Deban et al., 2011). PTX-3, a marker of the acute-phase inflammatory response, plays an important role in innate immunity and in modulation of the adaptive immune response (Yamasaki et al., 2009). Exercise-induced muscle damage is mainly diagnosed in two ways. The first way is the visualization methods, the other one is the observing the rates of enzyme activities exclusive to the muscle and levels of pro-inflammatory biomarkers in the plasma (Bazgir et al., 2015). In this study, we evaluated levels and the correlation of PTX-3, IL-6, and CRP following acute and chronic exercising in rats.
MATERIALS AND METHODS
Animals and experimental conditions
Twenty-six Wistar Albino 2-month-old male rats (200–250 g) were obtained from the Experimental Research Unit of the University of Pamukkale (Denizli, Turkey). They were reared under the supervision of a veterinarian, kept in well ventilated noises environment and allowed free axes to food and water. They were maintained on a 12/12 hr light–dark cycle under controlled temperature. All protocols used in this study were approved by the Pamukkale University Ethics Committee on animal research (PAUHDEK-2012/035).
Experimental design
The male animals were selected randomly and divided into three experimental groups: control (n=10), acute (n=7), and chronic (n=9). The control group was not trained (sedentary). Acute exercise group; with 0.1% incline on the treadmill, 3 days/week, 10 min/day, 20 m/min was run. The rats in chronic exercise group; with 20% incline on the treadmill, for 4 weeks, 7 days/week, 60 min/day, 0.1 m/min was run. All of the animals were exercised at the same time of every day.
Blood samples and measurements
At the end of the experimental period, all the animals were anesthetized with ketamin/xylazine HCl (75 mg/kg/10 mg/kg intraperitoneally). Blood samples were collected in heparinized tubes from the abdominal aorta of rats. Plasma samples were separated from cells by centrifugation at 3,000 rpm for 10 min. and were stored at −80°C until analysis. The plasma PTX-3, IL-6, and CRP concentrations were measured by an enzyme-linked immunosorbent assay (ELISA) method using an rat ELISA kit (SunRed Biotechnology Company, eBioscience An Affymetrix Company) in a multiplate ELISA reader (das, Digital and Analog Systems, Vimercate, MI, Italy).
Statistical analysis
Data was analyzed by IBM SPSS Statistics ver. 21.0 (IBM Co., Armonk, NY, USA). Continuous variables were expressed as mean±standard deviation and categorical variables as number and percent. Kruskal–Wallis variance analysis was used to compare more than two independent groups. Mann–Whitney U-test was used to compare two independent groups.
RESULTS
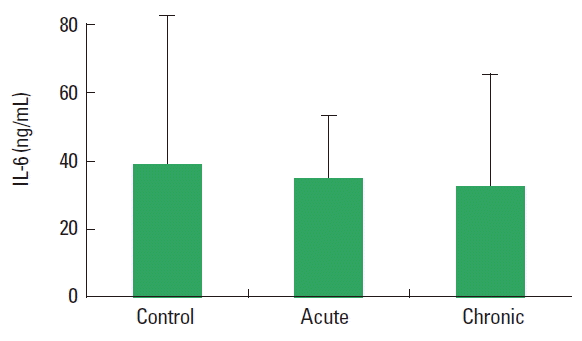
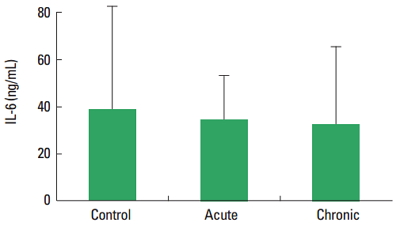
In current study, the PTX-3, IL-6, and CRP level was not observed statistically significant difference among control, acute, and chronic groups. There are not significantly different the concentration of IL-6 and CRP between acute and chronic exercise group (Figs. 1, 2) (P>0.05). The plasma levels of PTX-3 were found significantly increased in chronic exercise group compared to acute exercise group animals (Fig. 3) (P<0.05).
DISCUSSION
According to our results, the IL-6 level was not changed in acute and chronic exercise groups compared to sedentary group. An effective physical activity induces a series of changes in the immune system (Rosa Neto et al., 2009). It is known that the production of free oxygen radicals increased in the muscles and plasma during and following a highly severe contractile activity leads to muscle damage (Roebuck, 1999). The acute phase response activates the production of oxygen radicals together with various cytokines like interleukins and tumor necrosis factor-α. The most prominent cytokine produced during the contractions is IL-6 which is expressed within the muscle cells and released into the blood (Nielsen and Pedersen, 2008). Fischer et al. (2004) reported that reduced IL-6 mRNA response to exercise in trained human quadriceps muscle. Increased the IL-6 levels in both soleus and extensor digitorum longus muscles were reported in another study. Elevated IL-6 levels 48 hr after the last exercise session of the 8-week training period. The authors think that elevated muscular levels of IL-6 may be due to regeneration after injury (Isanejad et al., 2015). According to another study, plasma IL-6 concentration increases during different bouts of acute exercise (Pedersen and Febbraio, 2008). Kılıç et al. (2014) investigated that the possible role of moderate and strenuous swimming training on plasma and cerebrospinal fluid IL-6 levels during recovery from exhaustive exercise in rats. As a result of that study, plasma IL-6 levels were found unaltered among all groups. They conclude that exercise training intensity is an important factor determining IL-6 concentration after exhaustive exercise. In addition other researcher show that IL-6 expression was not affected by the training protocol (Lira et al., 2009). Mendham et al. (2011) demonstrated that the muscle damage response was notably augmented in the moderate-vigorous protocol compared to the low-intensity aerobic exercise, moderate-vigorous and low-intensity full-body resistance session protocols and this study evaluated the magnitude of the IL-6 response in light of the muscle damage response. The authors suggest that muscle damage may not be a major direct determinate of acute IL-6 release during exercise. Exercise training studies have reported equivocal results (decreases or no changes) in baseline levels of IL-6 (Nicklas et al., 2008). The results of previous studies investigating effects of muscle damage on IL-6 demonstrated conflicting results mainly depending on the type, duration, intensity of the exercise as well as the age at onset of exercise.
Plasma CRP concentrations in our study did not discriminate among all groups. Acute phase protein CRP have previously been reported to be associated with exercise-induced muscle damage, with evidence linking the stress and intensity of exercise to the subsequent magnitude of response of these markers within the circulation (Neubauer et al., 2008). Exercise training studies have uncertain results in levels of CRP (Donges et al., 2010). To date few studies report the acute exercise-induced CRP responses across different exercise modes and intensities within a sedentary, middle-aged cohort. Release of CRP is induced by IL-6 and responsible for recognition and clearance of damaged cells. (Neubauer et al., 2008) The present study indicates CRP to increase 24-hr postexercise within the higher-intensity protocols more so than the lower intensity protocols. Additionally, studies reporting the acute exercise-induced CRP response within a sedentary population are minimal; with previous training studies reporting a lack of association between IL-6 and CRP (Donges et al., 2010; Nicklas et al., 2008).
A novel biomarker, PTX-3, primarily released by adipocytes, monocytes/macrophages (Savchenko et al., 2008) and neutrophils (Jaillon et al., 2007). PTX-3 is produced rapidly in damaged tissue and may reflect more of a tissue-specific inflammatory response that includes smooth and skeletal muscle and adipocytes (Introna et al., 1996). Nakajima et al., (2010) demonstrated that acute moderate and intense aerobic exercise elevates PTX-3 concentrations in untrained individuals. Decreased PTX-3 levels during 6 months of cardiac rehabilitation exercise was reported (Fukuda et al., 2012). On the contrary, another study show that increased PTX-3 levels after an acute bout of maximal aerobic and resistance exercise, which may have been the result of exercise induced muscle damage. Also plasma levels of PTX-3 are elevated in endurance trained individuals compared to controls. Miyaki et al. (2011, 2012) found that endurance trained males exhibit elevated baseline PTX-3 plasma levels compared to sedentary controls while 8 weeks of aerobic training increased both VO2 peak and resting plasma PTX-3 levels in elderly women. Therefore, increased PTX-3 release from the potentially damaged muscle due to acute maximal exercise would not be surprising. In our present study the levels of PTX-3 was found to be higher in the chronic group compared to the acute group. The differences in the results of the studies may be due to the type of exercise applied and the time of application.
We can say that accorting to our results, response to muscle damage of IL-6 and CRP not affected by acute and chronic exercise. Plasma PTX-3 was found significantly higher in the chronic group than acute group but not control animals. Because of this reason we think that PTX-3 may be an inflammatory marker of muscle damage in chronic exercise or it may be increased to protects against muscle damage in chronic exercise. Also there is need to further studies for clearing this subject.