Size dependent classification of heat shock proteins: a mini-review
Article information
Abstract
Molecular chaperones are ubiquitous and abundant within cellular environments, functioning as a defense mechanism against outer environment. The range of molecular chaperones varies from 10 to over 100 kDa. Depending on the size, the specific locations and physiological roles of molecular chaperones vary within the cell. Multifunctionality of heat shock proteins (HSPs) expressed via various cyto-stress including heat shock have been spotlighted as a reliable prognostic target biomarker for therapeutic purpose in neuromuscular disease or cancer related studies. HSP also plays a critical role in the maintenance of proteins and cellular homeostasis in exercise-induced adaptation. Such various functions of HSPs give scientists insights into intracellular protective mechanisms in the living body thus HSPs can be target molecules to know the defense mechanism in cellular environment. Based on experimental results regarding small to large scaled HSPs, this review aims to provide updated important information regarding the modality of responses of intracellular HSPs towards extracellular stimulations. Further, the expressive mechanisms of HSPs data from tremendous in vivo and in vitro studies underlying the enhancement of the functionality of living body will be discussed.
INTRODUCTION
Heat shock proteins (HSPs) are a group of proteins that repress the denaturation of molecules by various stressful circumstances such as exercise, gravity, heat, oxygen, Ca2+ etc. HSPs are not limited to specific substrates. HSP family has a structurally common chaperone subset controlling the form of proteins’ folding (misfolding, facilitating, or reconstructuring), which maintains homeostasis of proteins to stressful circumstances (Dimauro et al., 2016). Heat-induced heat shock factor 1 controls transcriptional upregulation of HSPs (Brinkmeier and Ohlendieck, 2014). Their locations and functions vary within the cell. Small HSP (sHSP), HSP70, and HSP90 exist in the cytoplasm and glucose regulated proteins (GRP) 78 (a homologue of HSP70), Bip, HSP47 are located in mitochondria to transmit each of specific functional proteins to mitochondria. TCP1 (cytoskeleton forming chaperones) is associated with neogenesis of actin and tubulin and sHSP are involved in intracellular dynamics (polymerization and depolymerization). A transcript (unfolded Protein Response Element, URE; CCAAAN9 CCACG) binds to the sequence of the chaperone gene.
HSP families are made up by range from 10 to more than 100 kDa in molecular size, and are located in various cellular compartments. The expressional locations of homologue or cognate members of HSPs (such as HSC70 vs HSP70 or HSP90 α vs HSP90 β) are arbitrary according to the accumulated evidences indicating cell or tissue restricted expressions in vivo systems. Accumulated studies show that HSP families are located in various sites within cell; HSP10, HSP60, and HSP75 are located in mitochondria, however, others are present in the cytoplasm, cytosol, endoplasmic reticulum and nucleus in physiological conditions (Xu, 2002).
This review provides the role of each classified HSP. The following general description involving molecular chaperones shows the outline regarding the maintenance of physiological homeostasis of living body. Based on comprehensive studies of HSPs, this review offers important information regarding the mechanism related with the characteristic procedure of signal pathway according to the specifically cared substrate of each HSP, structural characteristics, specifically expressed location and those roles.
HSP 10–30
Molecular weight of sHSP varied from 15 to 43 kDa are also called heat shock protein β (HSPBs) and have a chaperoning function in the process of embryonic development. HSPBs are also interacting with cytoskeleton proteins to maintain the homeostasis of cytoskeleton proteins by preventing those from damages, which results in conservation of the cell function. HSPB1, HSPB6, HSPB7, and HSPB12 are involved in the development of respiratory morphology such as cardiac muscles and the lethal myopathy in diaphragm and cardiomyopathy is caused with the lack of those HSPBs (Juo et al., 2016; Ke et al., 2011; Rosenfeld et al., 2013).
HSP10 (chaperonin) is considered as a suppressor of maternal immune response via releasing from fetal placental unit (Noonan et al., 1979). This includes 101 amino acids and is used as a plausible biomarker in endometrial cancer (Dubé et al., 2007). HSP 10 in mitochondria plays a role in protein folding supplied by adenosine triphosphate (ATP) (Table 1) (Meyer et al., 2003). It has been reported that HSP10 is a by-product during the process of neoplastic cell proliferation and is considered as a growth factor in the cell (Quinn and Morton, 1992). HSP10 is also known as an obligatory autocrine growth factor in tumor cells (Quinn et al., 1990).
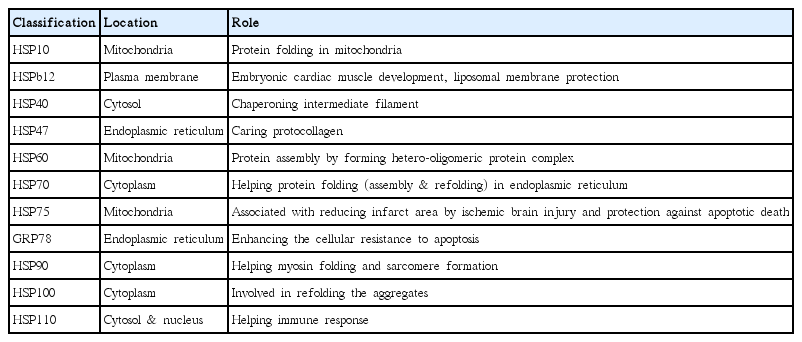
The location and role of HSP families
Molecular size 15–30 kDa HSPs are related with sHSP and it has been know that there are 9 sHSP in mammals (Mounier and Arrigo, 2002): Hsp27 (denoted Hsp25 in mice), αA- and αB-crystallins, Hsp20, HspB2, HspB3, cvHsp or HspB7, Hsp22 or HspB8, and HspB9. Human sHSP has 105–205 amino acids and α-crystallin domain as a homologous 80 sequenced residues (Ingolia and Craig, 1982). This domain shows highly conserved structure and 38%–60% of amino acid identity (Mounier and Arrigo, 2002). Alpha-crystallin structure is influenced by factors such as pH, temperature, calcium ions, and ionic strength (Mounier and Arrigo, 2002).
Each of the sHSP is phosphorylated via specific kinase to involve teleological signal pathway (Fig. 1). HSP16.5 has been structurally studied that it has dimeric α-crystallin domain as a basic structural domain unit and this common domain is possibly related with the basic function as a molecular chaperone (Bertz et al., 2010).
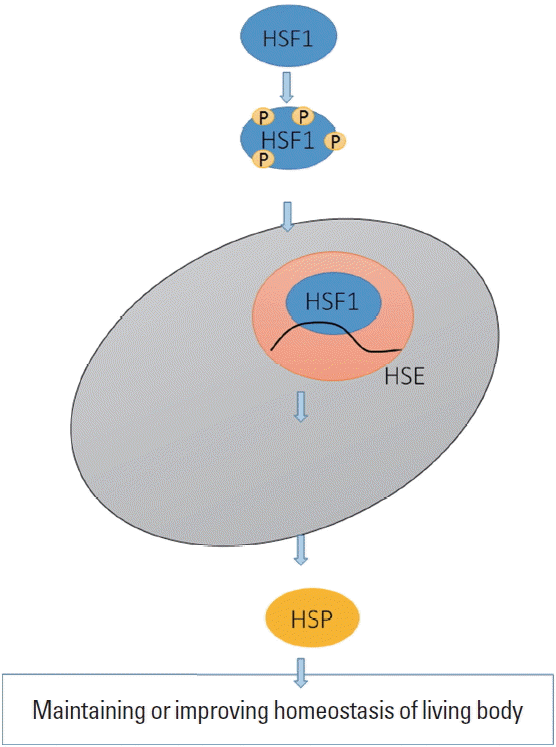
Heat shock protein (HSP) production. Heat shock factor 1 (HSF1) is changed to the activated form by phosphorylation and this translocates into nucleus to bind heat shock elements (HSEs). This process yields various types of HSPs to maintain or improve the homeostasis of living body.
Melkani et al. (2006) reported a function of sHSP on myosin enzymatic activity that αB-crystallin cares ATPase activity of myosin and the comparing rate shows 58% in relation to control (8%, without αB-crystallin) under 43°C for 30 min. Alpha B-crystallin maintaining ATPase activity of myosin prevent aggregation of the protein and this is probably related with myofibrilogenesis without myosin unfolding that induces muscular performance under stress condition or vigorous exercise (Melkani et al., 2006).
HSP 40–60
HSP40 locates in cytosol and cochaperones for intermediate filament with HSP70. Forty-three kDa sized HSPs has various amino acidic sequences that results in different N-terminal length. Those HSPs also comprise conserved α-crystallin core domains and C-terminal extension domain including highly conserved I-X-I/V motif (Ghosh et al., 2005).
HSP47 acting as a chaperone for procollagen has been also found to be involved in atherosclerosis. Strong focal expression of HSP47 is evident in atherosclerotic arteries of the collagenous regions. All cells expressing type I procollagen also express HSP47. Heat shock and oxidized low density lipoprotein stimulate the expression of HSP47 mRNA in smooth muscle cells. These findings identify HSP47 as a novel constituent of human coronary atheroma, and selective upregulation by stress raises the possibility that HSP47 may be a determinant of plaque stability. Collagen, an extracellular matrix substrate, requires HSP47 to be transported within the cell and folded from three helices. HSP47 located in endoplasmic reticulum and functions to support collagen folding and its release to extracellular environments.
HSP56 is located in the cytosol. HSP60 forms a large (970 kDa) hetero-oligomeric protein complex called the TCP1 ring complex (containing TCP1 and several other proteins), which is essential for protein assembly.
The HSP60 family has been shown to be involved in the development of many diseases, such as adjuvant arthritis in rats, rheumatoid arthritis in humans, insulin-dependent diabetes mellitus in mice, and systemic sclerosis in humans. HSP60 is located in mitochondria.
HSP 70–100
HSP70 is in the cytosol and has family of GRP78 which plays role in helping protein folding assembly and refolding, transporting and blocking protein degradation in endoplasmic reticulum. HSP70 is expressed by physical exercise (Banfi et al., 2004; McClung et al., 2008; Ogawa et al., 2011; Shastry et al., 2002).
HSP70 and HSP90 expression levels are variant according to different individual athletic abilities and seem to be upregulated in response to heat acclimation (Banfi et al., 2004; McClung et al., 2008; Shastry et al., 2002).
As a chaperone, HSP70 plays a role in the assembly and transport of newly synthesized proteins within cells, as well as in the removal of denatured proteins. HSP70 cares matured proteins to be folded for those functional structures. The modification of carbohydrate is involved in GRP78 and Bip. GRP78 cares matured proteins to be folded for those functional structures. GRP78 has a resistance to the ultraviolet rays and cisplatin (anticancer drug). HSP90 is located in the cytosol and nucleus. GRP94 is in the endoplasmic reticulum.
Downhill (eccentric contraction exercise induced muscle damage by increased delayed onset muscle soreness) in hot conditions elevates the largest expression of HSP72 and HSP90 α mRNA level (Tuttle et al., 2015).
In related with myocyte formation, HSP90 α plays a key role in myosin folding and sarcomere formation (Du et al., 2008; Etard et al., 2007; Hawkins et al., 2008; Srikakulam et al., 2008).
HSP90 consists of N-terminal ATP binding domain, substrate interacting middle domain, and C-terminal dimerization domain (Jackson, 2013). By ATP binding to the N-terminal ATP-binding domain, HSP90 regulates myosin thick filament formation and muscle myofibrilogenesis (Hawkins et al., 2008).
HSP90 binds steroid receptors, protein kinases, intermediate filaments, microtubules, and actin microfilaments in a specific manner. HSP90 is an essential component of the glucocorticoid receptor, assembled in a complex of several proteins. HSP90 is divided into HSP90 α and HSP90 β. HSP40 and HSP70 are cochaperones with the HSP90s. HSP90s are located in cytoplasm and endoplasmic reticulum to play roles in folding newly made proteins.
HSP 100~
HSP100 is located in the cytoplasm and cochaperones with HSP40, HSP70, and HSP90. HSP100 plays a role in refolding the aggregate.
HSP110 is in the cytosol and nucleus (Dimauro et al., 2016). HSP110 and HSP70 super family commonly have the presence of a loop structure and HSP110 helps immune response (Zuo et al., 2016). HSP110 is also in working with HSP70 or GRP78 to fold proteins and counter stress for cell survival (Gething and Sambrook, 1992; Hartl, 1996).
Zuo et al. (2016) vaccinated with purified HSP110 results in reduced cancer metastasis thus Large HSPs amplify inflammatory signals in the cellular environment, suggesting potential endogenous immuno-stimulation during injury such as infection (Zuo et al., 2016).
There are comparatively very rare studies regarding large HSPs especially in the effect of exercise on the expression of large HSP and its function.
CONCLUSIONS
Over several decades, HSPs have been known as a potent factor playing a role in maintaining the homeostasis of living body. Versatile functions of HSPs according to those wide ranges of molecular weight that care those each substrate have been increasingly attentive with other research subjects such as exercise, carcinogenesis, muscles, etc. The broad ranges of HSPs are organelle-specifically expressed within the body to play physiological roles via interacting with various signal pathways. Provoking these pathways in a positive way (e.g., physical exercise) enhance HSPs signal pathway and maintain/improve vital function of HSPs in the living body.
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.