Treadmill exercise in obese maternal rats during pregnancy improves short-term memory through neurogenesis in the hippocampus of rat pups
Article information

Abstract
Maternal obesity is known to increase the likelihood of offspring becoming obese, high blood pressure, and other metabolic disorders. After inducing obesity, the effect of treadmill exercise in maternal rats during pregnancy on short-term memory was investigated in relation to neurogenesis in rat pups. Short-term memory was declined in rat pups born to obese maternal rats, and treadmill running during pregnancy alleviated short-term memory impairment in rat pups born to obese maternal rats. The number of doublecortin (DCX)-positive and 5-bro-mo-2′-deoxyuridine (BrdU)-positive cells in the hippocampal dentate gyrus was decreased in rat pups born to obese maternal rats. Treadmill running during pregnancy increased the number of DCX-positive and BrdU-positive cells in rat pups born to obese maternal rats. Expression of brain-derived neurotrophic factor (BDNF) and tyrosine kinase B (TrkB) in the hippocampus was decreased in the rat pups born to obese maternal rats. Treadmill running during pregnancy increased the expressions of BDNF and TrkB in rat pups born to obese maternal rats. Enhancing effect of short-term memory by treadmill exercise may be due to increased neurogenesis through activation of the BDNF-TrkB signaling pathway by treadmill exercise.
INTRODUCTION
Maternal health during pregnancy refers to the health of the fetus, and maternal obesity is known to have a direct effect on the induction of diabetes in children (Tozuka et al., 2010). Maternal obesity is known to increase the likelihood of offspring becoming obese, high blood pressure, and other metabolic disorders (Chang et al., 2008; Shankar et al., 2008). Reduced brain-derived neurotrophic factor (BDNF) expression, impaired spatial cognitive function, and suppressed neurogenesis in the hippocampus have been observed in pups born to obese maternal rats (Tozuka et al., 2010). The hippocampus is a component of the brain limbic system and the hippocampus incorporates all information such as short-term memory, long-term memory, spatial learning memory, and spatial working memory (Snyder et al., 2005).
Various neuronal markers appear on developing neurons. Immunohistochemistry for 5-bromo-2′-deoxyuridine (BrdU) is used for quantification of new cell generation (Park et al., 2019), and the neuronal nuclear (NeuN) antigen is considered as a useful marker for neuronal maturation (Park et al., 2019). Doublecortin (DCX) is a microtubule-related protein and specifically expressed during cell production in the central nervous system (Couillard-Despres et al., 2001). DCX expression refers a marker of neuronal precursor cells and indicates the migration and development of neurons (Park et al., 2019).
BDNF regulates neuronal development, differentiation, and survival through the receptor tyrosine kinase B (TrkB), and BDNF is secreted by activity-dependent modulators (Massa et al., 2010). Learning and memory skills in rat pups were increased with exercise of maternal rats during pregnancy (Akhavan et al., 2008), and low-intensity exercise of maternal rats during pregnancy was associate with improved memory function of pups born to maternal rats with posttraumatic stress disorder (Seo et al., 2013).
There are insufficient scientific evidences for the effect of exercise on short-term memory in children during pregnancy in obese mothers. In this study, step-down avoidance task was performed to assess the impact of treadmill exercise in obese maternal rats during pregnancy on the short-term memory of rat pups. The expression of BrdU and DCX was detected by immunohistochemistry and the expression of BDNF and TrkB, was measured by western blot analysis.
MATERIALS AND METHODS
Animals
Approval number was achieved from the Kyung Hee University Institutional Animal Care and Use Committee in Seoul, Korea (KHUASP [SE]-17-099). The experimental animals used in this study were 4-week-old Sprague-Dawley male rats (n=8) and female rats (n=16). In order to induce obesity, female rats were supplied with the high-fat diet (60% fat) purchased from Central Laboratory Animals (Seoul, Korea). Female rats were classified into the normal diet group (n=8) and the high-fat diet group (n=8) for a total of 7 weeks, including gestation period and lactation period. Female rats were bred individually 48 hr after mating with male rats. Female rats with confirmed pregnancy were divided into normal diet group, normal diet and treadmill exercise group, high-fat diet group, high-fat diet and treadmill exercise group. The born rat pups were divided into control group, treadmill exercise group, high-fat diet group, high-fat diet and treadmill exercise group (n=10), according to the classification status of maternal rats. All experiments were conducted in compliance with the recommendations of the National Institutes of Health and the Korean Academy of Medical Sciences.
Treadmill exercise protocol and BrdU injection
The running speed of treadmill during the pregnancy was 3 m/min for the first 5 min, 5 m/min for the next 5 min, 8 m/min for the last 20 min, 1 time for 30 min, 5 times a week, and for 4 weeks. BrdU (Sigma Chemical Co., St. Louis, MO, USA) was injected intraperitoneally with 50 mg/kg once a day for 5 days from the 15th day of pregnancy 60 min before the treadmill exercise.
Step-down avoidance task
In the same manner, as described below (Ko et al., 2019), the latency in the step-down avoidance task was measured to assess short-term memory. The rat pups were trained with a step-down avoidance task after last treadmill exercise. The rat pups were placed 7×25×2.5-cm platform and the platform faced a 42×25-cm grid of parallel 0.1-cm caliber stainless-steel bars spaced 1 cm apart. In training session, the animals were subjected to a 0.5-mA scramble foot shock for 2 sec as soon as they descended. One hour after training, the latency (in seconds) of each animal was measured. We defined the latency as the time interval between when the rat descends and puts all four feet on the grid, and a latency of more than 300 sec was calculated as 300 sec.
Tissue preparation
Immediately after the Morris water maze task, the rat pups were anesthetized with intraperitoneal injection of Zoletil 50 (10 mg/kg, Vibac Laboratories, Carros, France). The rat pups were cardiac perfused with 50-mM phosphate-buffered saline (PBS) and then fixed by 4% paraformaldehyde dissolved in 100-mM phosphate buffer (PB; pH, 7.4). After the brains were removed, the brains were postfixed overnight using the same fixative and transferred to a 30% sucrose solution to prevent freezing. The 40-μm thick sagittal sections were created using a freezing microtome (Leica, Nussloch, Germany).
Immunohistochemistry for DCX
In the same manner, as described below, immunohistochemistry for DCX was performed (Park et al., 2019). The sections were treated with PBS for 10 min and then treated again with 1% H2O2 for 20 min. The sections were treated with goat anti-DCX antibody (1:1,000; Oncogene Research Product, Cambridge, UK) for 2 hr. The sections were treated with the biotinylated goat secondary antibody (1:500; Vector Laboratories, Burlingame, CA, USA) for another 1 hr, washed, and treated with ABC complex (1:100; Vector Laboratories). Using the 0.03% diaminobenzidine, labeling was visualized and the sections were mounted onto gelatin-coated slides. After air drying the slides at room temperature overnight, the coverslips were mounted using Permount (Thermo Fisher Scientific Inc., Waltham, MA, USA).
Immunohistochemistry for BrdU
In the same manner, as described below, immunohistochemistry for BrdU was performed (Park et al., 2019). The sections were first permeabilized by treating with 0.5% Triton X-100 in PBS for 20 min, then treated with 50% formamide-2 X standard saline citrate at 65°C for 2 hr, denatured in 2 N HCl at 37°C for 30 min, and washed twice in 100-mM sodium borate (pH, 8.5). The sections were treated with BrdU-specific mouse monoclonal antibody (1:600; Roche, Mannheim, Germany) at 4°C overnight. The sections were treated with biotinylated mouse secondary antibody (1:200; Vector Laboratories) for 1 hr and treated with ABC complex (1:100; Vector Laboratories) for 1 hr. To visualized, the sections were treated for 5 min in 50-mM Tris-HCl (pH, 7.6) containing 0.03% diaminobenzidine, 0.03% H2O2 and 40-mg/mL nickel chloride.
To differentiate neurons, mouse antineuronal nuclei antibody (1:1,000; Chemicon International, Temecula, CA, USA) was used in the same sections after BrdU labeling. The sections were treated with a biotinylated anti-mouse secondary antibody for l hr. For staining, the sections were treated in a reaction mixture consisting of 0.03% diaminobenzidine and 0.03% H2O2 for 5 min. The sections were mounted onto gelatin-coated slides, air-dried overnight at room temperature, and coverslips were mounted using Permount (Thermo Fisher Scientific Inc.).
Western blot analysis
In the same manner, as described below, western blotting for the determination of BDNF and TrkB was conducted (Park et al., 2019). Hippocampal tissues were homogenized on ice and lysed in a lysis buffer consisting of 1-mM phenylmethylsulfonyl fluoride, 150-mM NaCl, 0.1% sodium dodecyl sulfate (SDS), 0.5% deoxycholic acid, 1% Nonidet P40, 100-mg/mL leupeptin, 50-mM Tris-HCl (pH, 7.5). Colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA) was used to measure protein content. Total protein 30 μg was separated on SDS-poly-acrylamide gels and transferred to a nitrocellulose membrane, the membrane was blocked with dehydrated milk and then treated with primary antibodies. Mouse anti-β actin (1:3,000; Santa Cruz Biotechnology, CA, USA), rabbit anti-BDNF (1:1,000; Santa Cruz Biotechnology), and rabbit anti-TrkB (1:1,000; Santa Cruz Biotechnology) were used as the primary antibody.
After washing, appropriate horseradish peroxidase-conjugated secondary antibodies were used and incubation was performed at room temperature. Enhanced chemiluminescence detection system (Santa Cruz Biotechnology) was used to measure the expression of bands, and bands were quantified using an Image-Pro Plus computer-assisted image analysis system (Media Cyberbetics Inc., Silver Spring, MD, USA).
Data analysis
IBM SPSS Statistics ver. 21.0 (IBM Co., Armonk, NY, USA) was used to analyze the data, and one-way analysis of variance using the Duncan post hoc test was used for comparisons between groups. The results were presented as the mean±standard error of the mean, and P<0.05 was shown to be statistically significant.
RESULTS
Short-term memory
In order to investigate the effect of treadmill exercise during pregnancy of obese maternal rats on the short-term memory in rat pups, a step-down avoidance task was performed (Fig. 1). In the high-fat diet group, the latency was significantly reduced compared with the control group (P<0.05). Treadmill running was found to significantly increase the latency in the high-fat diet group (P<0.05).

Short-term memory of the rat pups. A, control group; B, treadmill exercise group; C, high-fat diet group; D, high-fat diet and treadmill exercise group. *P<0.05 compared with the control group. #P<0.05 compared with the high-fat diet group. Data represents the mean±standard error of the mean.
Number of DCX-positive cells
In order to investigate the effect of treadmill exercise during pregnancy of obese maternal rats on the neuronal production in hippocampal dentate gyrus of rat pups, the number of DCX-positive cells was observed (Fig. 2). The number of DCX-positive cells was significantly reduced compared with the control group (P< 0.05). Treadmill running was found to significantly increase the number of DCX-positive cells in the high-fat diet group (P<0.05).
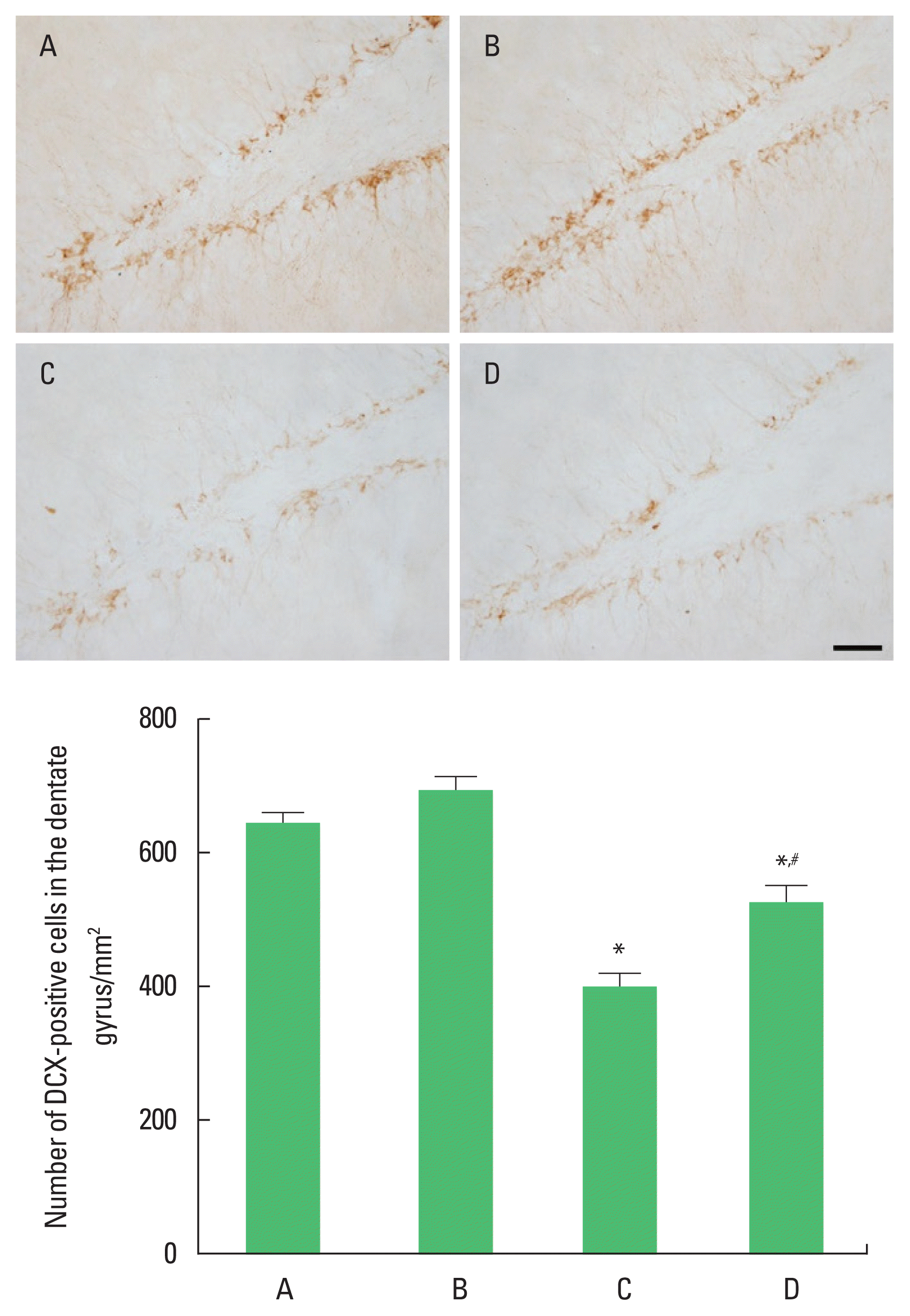
Doublecortin (DCX) expression in the hippocampal dentate gyrus of rat pups. Upper panel: photomicrographs of DCX-positive cells. The scale bar represents 50 μm. Lower panel: number of DCX-positive cells in each group. A, control group; B, treadmill exercise group; C, high-fat diet group; D, high-fat diet and treadmill exercise group. *P<0.05 compared with the control group. #P<0.05 compared with the high-fat diet group. Data represents the mean± standard error of the mean.
Number of BrdU-positive cells
In order to investigate the effect of treadmill exercise during pregnancy of obese maternal rats on the cell proliferation in hippocampal dentate gyrus of rat pups, the number of BrdU-positive cells was observed (Fig. 3). The number of BrdU-positive cells was significantly reduced compared with the control group (P< 0.05). Treadmill running was found to significantly increase the number of BrdU-positive cells in the high-fat diet group (P<0.05).
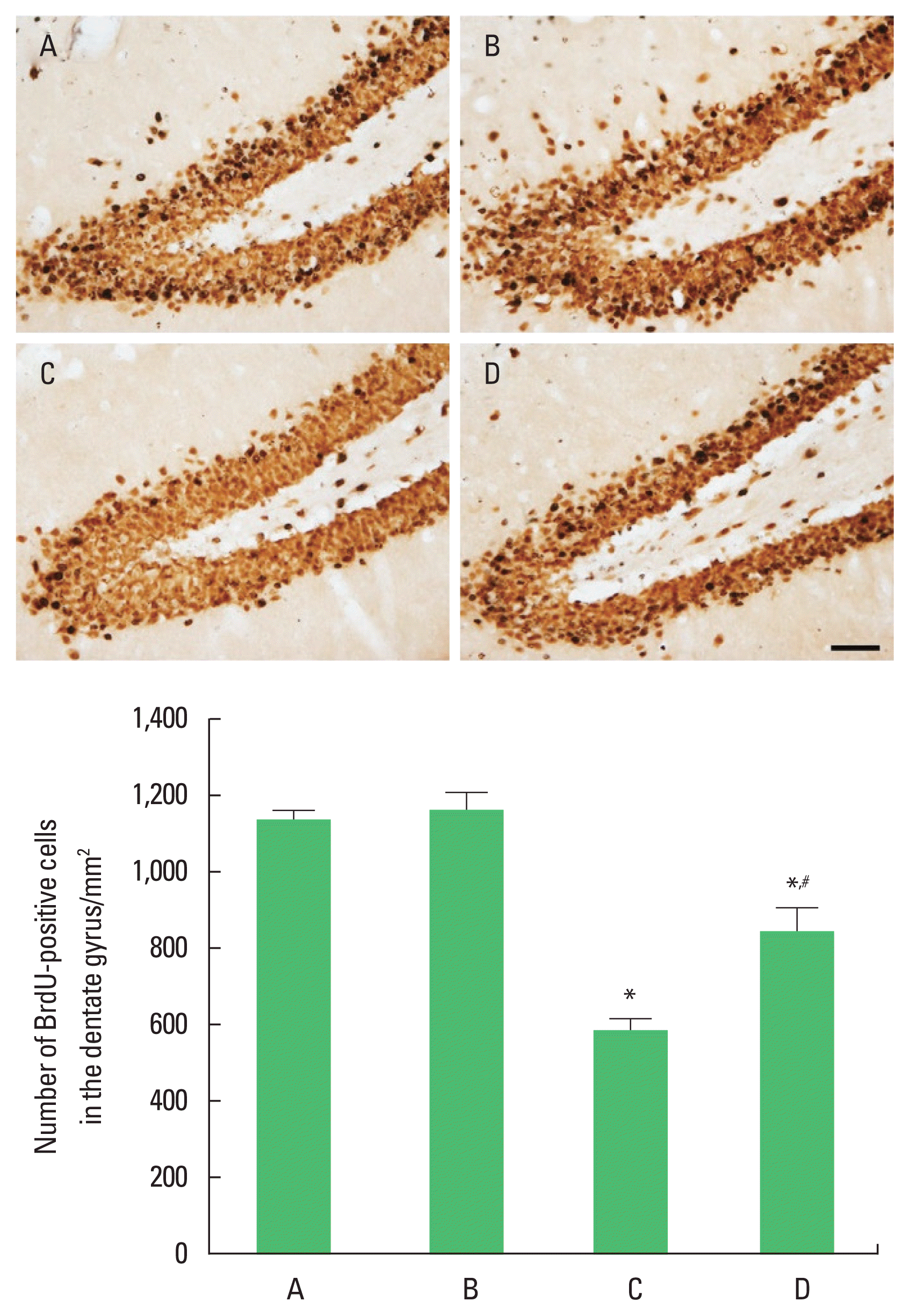
Cell proliferation in the hippocampal dentate gyrus of rat pups. Upper panel: photomicrographs of 5-bromo-2′-deoxyuridine (BrdU)-positive cells. The scale bar represents 50 μm. Lower panel: number of BrdU-positive cells in each group. A, control group; B, treadmill exercise group; C, high-fat diet group; D, high-fat diet and treadmill exercise group. *P<0.05 compared with the control group. #P<0.05 compared with the high-fat diet group. Data represents the mean±standard error of the mean.
BDNF and TrkB expression
In order to investigate the effect of treadmill exercise during pregnancy of obese maternal rats on the expression of BDNF and TrkB in the hippocampus of rat pups, the expression of BDNF and TrkB was observed (Fig. 4). The expression of BDNF and TrkB was significantly reduced in the high-fat diet group compared with the control group (P<0.05). Treadmill running was found to significantly increase the expression of BDNF and TrkB in the high-fat diet group (P<0.05).

Brain-derived neurotrophic factor (BDNF) and tyrosine kinase B (TrkB) expression in the hippocampus of rat pups. Upper panel: western blot analysis of BDNF and TrkB in the hippocampus. Left lower panel: optical density of BDNF expression in each group. Right lower panel: optical density of TrkB expression in each group. A, control group; B, treadmill exercise group; C, high-fat diet group; D, high-fat diet and treadmill exercise group. *P<0.05 compared with the control group. #P<0.05 compared with the high-fat diet group. Data represents the mean±standard error of the mean.
DISCUSSION
Ingestion of a high-fat diet during pregnancy enhances inflammatory cytokines, affecting the neurological development of offspring (Sullivan et al., 2014), and it has been reported that the offspring’s behavior becomes abnormal (Sullivan et al., 2010). During pregnancy and lactation, the mother’s intake of saturated transfat and high-fat diet decreases cognitive function (Bilbo and Tsang, 2010), and reduces neurotrophic factors associated with cognitive function in the hippocampus of the brain (Tozuka et al., 2010). On the contrary, exercise is well known as a representative factor that improves cognitive function, and it has been reported that exercise enhances learning ability and memory function by increasing the production of neurons in the hippocampus (Trejo et al., 2001). Mother’s exercise during pregnancy increases the offspring’s ability to learn and memory (Akhavan et al., 2008; Kim et al., 2007) and increases neurogenesis in the hippocampus of pups born under stress (Kim et al., 2013). In the results of current study, short-term memory was reduced in the high-fat diet group compared with the control group. Treadmill running enhanced short-term memory in the high-fat diet rat pups.
The hippocampus belongs to the limbic system of the brain and is an important brain part involved in the formation of learning and memory in relation to neurogenesis. It is also the site of the greatest neuroplasticity (Snyder et al., 2005). During pregnancy or lactation, maternal nutrition and hormones exert critical effect in fetal growth and neural circuit formation in the brain (Férézou-Viala et al., 2007). Swimming exercise in maternal rats during pregnancy improved short-term memory and neuronal production in offspring (Lee et al., 2006), and treadmill exercise of rats during pregnancy enhanced the brain function in offspring (Kim et al., 2007). In the results of current study, the number of DCX-positive cells and BrdU-positive cells in the hippocampus was decreased in the high-fat diet group compared with the control group. Treadmill running increased the number of DCX-positive cells and BrdU-positive cells in the high-fat diet rat pups.
BDNF is secreted from various brain regions and is an important representative substance for the growth and development of nerve cells and neuroplasticity. BDNF improves the survival of neurons by increasing resistance to nerve damage (Mizuno et al., 2000) and plays an important role in neuronal production in the hippocampus (Donovan et al., 2008). Ingestion of a high-fat diet reduced the production of neurons and expression of BDNF in the hippocampus (Stranahan et al., 2008), and long-term consumption of a high-fat diet impaired hippocampal function, which is responsible for learning and memory (Tozuka et al., 2010). BDNF regulates the development and differentiation of neurons by binding to the receptor TrkB, and BDNF is secreted by an activity-dependent manner (Bramham and Messaoudi, 2005). In the results of current study, the expression of BDNF and TrkB in the hippocampus was decreased in the high-fat diet group compared with the control group. Treadmill running enhanced BDNF and TrkB expression in the high-fat diet rat pups.
The current study revealed that neuronal cell generation and survival in the hippocampus of rat pups were increased through maternal treadmill exercise in the high-fat diet group. Treadmill exercise in the obese maternal rats increased BDNF and TrkB expression in offspring born to obese mothers, which has a positive effect on neuronal cell production and survival.
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.
ACKNOWLEDGMENTS
This work was supported by 2019 Sangmyung University Research Foundation of Korea.