Exercise before pregnancy exerts protective effect on prenatal stress-induced impairment of memory, neurogenesis, and mitochondrial function in offspring
Article information

Abstract
Stress during pregnancy has a negative effect on the fetus. However, maternal exercise has a positive effect on the cognitive function of the fetus and alleviates the negative effects of stress. This study aimed to demonstrate whether exercise before pregnancy has a protective effect on prenatal stress-induced impairment of memory, neurogenesis and mitochondrial function in mice offspring. In this experiment, immunohistochemistry, Western blot, measurement of mitochondria oxygen respiration, and behavior tests were performed. Spatial memory and short-term memory of the offspring from the prenatal stress with exercise were increased compared to the offspring from the prenatal stress. The numbers of doublecortin-positive and 5-bromo-2′-deoxyuridine-positive cells in the hippocampal dentate gyrus of the offspring from the prenatal stress with exercise were higher compared to the offspring from the prenatal stress. The expressions of brain-derived neurotrophic factor, postsynaptic density 95 kDa, and synaptophysin in the hippocampus of the offspring from the prenatal stress with exercise were enhanced compared to the offspring from the prenatal stress. Oxygen consumption of the offspring from the prenatal stress with exercise were higher compared to the offspring from the prenatal stress. Exercise before pregnancy alleviated prenatal stress-induced impairment of memory, neurogenesis, and mitochondrial function. Therefore, exercise before pregnancy may have a protective effect against prenatal stress of the offspring.
INTRODUCTION
Prenatal stress reduces cognitive function in infancy and adulthood (Polanska et al., 2017). Stress during pregnancy reduces mitochondrial content and increases oxidative stress in the hippocampus of offspring, contributing to reduced neurogenesis and cognitive decline (Zheng et al., 2015). The hippocampus plays a variety of roles, including controlling short-term and long-term memory and emotion-related information. Newly generated neurons enhance the learning and memory functions of the hippocampus (van Praag et al., 2005). Aging, irradiation, and stress reduce neurogenesis in the dentate gyrus of the hippocampus (Karten et al., 2005; Madsen et al., 2003).
Hippocampal mitochondria regulate calcium homeostasis and energy-requiring neurotransmission, mechanisms important for learning and memory (Mehdizadeh et al., 2017). Madrigal et al. (2001) reported that chronic stress inhibits the mitochondrial respiratory chain, resulting in mitochondrial dysfunction. Rezin et al. (2009) stated that the pathological causes of depression and bipolar disorder are mitochondrial dysfunction and biochemical damage to the mitochondrial electron transport system. Stress is known to reduce mitochondrial oxygen consumption and ATP production associated with neurogenesis (Seo et al., 2019).
Maternal exercise during pregnancy is known to bring about beneficial changes in the health status of the fetus and to reduce anxiety and depression (Murtezani et al., 2014). Maternal exercise may have beneficial effects on the cognitive development of offspring. For example, offspring of mothers who exercised prenatally have been known to show improved language skills. These findings may be related to brain changes, such as enhanced fetal brain maturation (Labonte-Lemoyne et al., 2017; Murtezani et al., 2014). Exercise before and during pregnancy is effective in improving brain function and development in offspring (Wiebe et al., 2015). Additionally, exercise before pregnancy may reduce changes caused by prenatal stress (Luft et al., 2020). Exercise has a protective effect against stress-induced brain changes (Bustamante et al., 2020; Park et al., 2020).
Several studies have demonstrated the beneficial effects of maternal exercise during pregnancy on the child’s neurobehavioral development and hippocampal neuroplasticity. However, the effects of maternal exercise before pregnancy on fetal stress-related cognitive function, hippocampal neurogenesis, neuroplasticity, and mitochondrial function are still unknown. Therefore, this study aimed to demonstrate whether exercise before pregnancy has a protective effect on prenatal stress-induced impairment of memory, neurogenesis and mitochondrial function in mice offspring. For this experiment, immunohistochemistry, western blot, mitochondrial oxygen respiration measurement, and behavioral tests were performed.
MATERIALS AND METHODS
Experiment animals
Four-week-old C57BL/6 mice were used in the experiments. Mice were individually housed in cages at a controlled temperature (23°C±2°C) and maintained under light-dark cycles comprising 12 hr each of light and dark. This study was approved by the Kyung Hee university Institutional Animal Care and Use Committee (Seoul, Korea) (KHUASP (SE)-21-179). Before pregnancy, female C57BL/6 mice were divided into exercise and nonexercise groups (n=8 in each group). After pregnancy, these mice were randomly divided into the following groups (n=4 per group): control group, exercise group, stress exposed during pregnancy group, and stress-exposed during pregnancy with exercise group. The offspring were assigned into the four groups (n=10 per group): offspring from the control group, offspring from the exercise group, offspring from the prenatal stress group, and offspring from the prenatal stress with exercise group.
Exercise protocol before pregnancy
According to previously described methods (Hong et al., 2020), the exercise group exercised on a treadmill made for animals for 6 days a week for 8 consecutive weeks before mating. The exercise load consisted of running at an initial speed of 3 m/min for the first 5 min, 5 m/min for the next 5 min, and 8 m/min for the last 20 min at an incline of 0°. Animals in the nonexercise group did not run but stayed on the treadmill for the same amount of time.
Stress exposure to pregnant mice
Pregnant mice were subjected to electric foot shocks by placing each mouse in a box with a bottom made of stainless steel grids, according to following method with minor modifications (Shi and Davis, 2002). A stimulator was used to deliver scrambled electric shocks through a floor grid. An interval timer was connected to the stimulator, which delivered 5-sec electric shocks every 30 sec for 5 min at an intensity of 0.5 mA.
Elevated plus maze test
Anxiety-related behaviors of pups were assessed using the elevated plus maze test according to following method (Park et al., 2019). The plus maze was constructed black acrylic with two open arms (50×10×36 cm) and two closed arms (50×10×36 cm), where the two arms met and connected to a central platform (10×10 cm). The entire plus maze was raised 60 cm above the floor and illuminated by a 100-W light bulb fixed 2 m above the maze floor. Mice were placed on the central platform facing a closed arm and allowed to explore the maze freely for 300 sec. We measured entry and time spent in the open arms were measured.
Morris water maze task
The Morris water maze task was used to assess spatial learning and working memory according to following method (Seo et al., 2019). One day before training, the pups were habituated to swimming for 60 sec in a pool without a platform. All pups were trained and recorded 3 times a day for five consecutive days. Probe trials were performed 24 hr after the final training session. When pups discovered the platform, it was allowed to remain there for 30 sec. If the pups did not find the platform within 60 sec, it was manually guided to the platform. Pups underwent a 60-sec retention probe test, after which the platform was removed from the pool. Data were collected automatically using the Smart Video Tracking System (Smart version 2.5, Panlab, Barcelona, Spain).
Step-through avoidance task
To assess short-term memory, we performed a step-through avoidance task 24 hr after training according to following method (Park et al., 2020). Latency was measured for each group. During training session, pups were placed in a light compartment by a lamp (60 W) and the guillotine door was raised to allow pups to enter the dark compartment. The guillotine door was closed when the pups’ hind legs entered the dark compartment. An electric foot shock (0.2 mA) was delivered for 2 sec through the grid floor. This process was repeated 24 hr after the training session. The time elapsed before the pups entered the dark compartment was recorded. Delays longer than 180 sec are considered 180 sec.
Doublecortin immunohistochemistry
Doublecortin (DCX) immunohistochemistry was performed according to following method (Kim et al., 2015). The sections were incubated overnight with mouse anti-DCX antibody (1:500; Santa Cruz Biotechnology, Santa Cruz, CA, USA), and then incubated for 1 hr with biotinylated mouse secondary antibody. The bound secondary antibody was amplified using a Vector Elite ABC kit (Vector Laboratories, Burlingame, CA, USA). The antibody–biotin–avidin-peroxidase complexes were visualized using 0.02% diaminobenzidine. The sections were mounted on gelatin-coated slides. The slides were air-dried overnight at room temperature, and the coverslips were mounted using Permount (Fisher Scientific, New Jersey, NJ, USA).
5-bromo-2′-deoxyuridine immunohistochemistry
5-bromo-2′-deoxyuridine (BrdU) immunohistochemistry was performed according to following method (Lee et al., 2016). The brain sections were first permeabilized by treating in 0.5% Triton X-100 in phosphate-buffered saline for 20 min, incubated in 50% formamide-2 x standard saline citrate at 65°C for 2 hr, denatured in 2 N HCl at 37°C for 30 min, and washed twice in 100 mM sodium borate (pH, 8.5). Tissue sections were treated overnight at 4°C with BrdU-specific mouse monoclonal antibody (1:600; Roche, Mannheim, Germany). The sections were then treated with biotinylated mouse secondary antibody (1:200; Vector Laboratories) for 1 hr. The sections were incubated in 50 mM Tris-HCl (pH 7.6) with 0.02% diaminobenzidine, 40-mg/mL nickel chloride, and 0.03% hydrogen peroxide for 5 min. After BrdU staining, differentiation of BrdU-positive cells was evaluated in the same section using a mouse antineuronal nuclei antibody (1:1,000; Chemicon International, Temecula, CA, USA). Slides were then air-dried overnight at room temperature, and coverslips were mounted using Permount (Fisher Scientific).
Western blotting for brain-derived neurotrophic factor, postsynaptic density 95 kDa, and synaptophysin
According to following method (Seo et al., 2019), the hippocampal tissues were collected, homogenized on ice, and lysed in a lysis buffer. Proteins were quantified using Bradford protein assay (Bio-Rad, Hercules, CA, USA). After electrophoresis, proteins were transferred onto nitrocellulose membrane (GE Healthcare Life Sciences, Chicago, IL, USA). The membrane was blocked with skim milk, then which was treated with mouse β-actin antibody (1:1,000; Santa Cruz Biotechnology), rabbit brain-derived neurotrophic factor (BDNF) antibody (1:1,000; Bioss Antibodies, Woburn, MA, USA), rabbit postsynaptic density 95 kDa (PSD-95) antibody (1:1,000; Abcam, Cambridge, UK), and rabbit synaptophysin (1:1,000; Abcam). Horseradish peroxidase-conjugated anti-mouse for β-actin (1:3,000) and anti-rabbit for BDNF, PDS-95, synaptophysin (1:5,000) were used as secondary antibodies.
Measurement of mitochondria oxygen respiration
High-resolution oxygen consumption measurements were performed using an Oroboros Oxygraph-2 K (Oroboros Instruments, Innsbruck, Austria) according to following method (Anderson et al., 2011). The sample was moved to a chamber with 2 mL of assay respiration buffer. The mitochondrial oxygen consumption protocol comprised 5 mM glutamate, 2 mM malate (complex I substrates), 4 mM adenosine diphosphate infusion (ADP) (state 3 condition), and 3 mM succinate. The oxygen consumption rate was represented as pmol/min/mg of wet tissue weight.
Data analysis
The detected bands were quantified using a computer-assisted Image-Pro Plus analysis system (Media Cybernetics, Inc., Silver Spring, MD, USA) to compare the relative expression levels. All data were analyzed using IBM SPSS Statistics ver. 25.0 (IBM Co., Armonk, NY, USA) and expressed as mean±standard error of the mean. One-way analysis of variance followed by Duncan post hoc test was used to compare the groups. Statistical significance was set at P<0.05.
RESULTS
Behavioral tests
The results of behavioral tests are shown in Fig. 1. In the Morris water maze task, the proportion of time taken by the offspring exposed to prenatal stress in occupying the probe quadrant was lower than that required by those not exposed to prenatal stress. Offspring from the prenatal stress with maternal exercise group spent a longer time occupying the probe quadrant compared to the time spent by offspring from the prenatal stress group. In the elevated plus maze task, the offspring exposed to prenatal stress spent less time in the open arm than those not exposed to prenatal stress did. Offspring belonging to the prenatal stress with maternal exercise group spent more time in the open arms compared to that spent by their counterparts from the prenatal stress group. The offspring exposed to prenatal stress entered the open arms fewer times than those not exposed to prenatal stress did. Offspring from the prenatal stress with maternal exercise group entered the open arms more times than the offspring from the prenatal stress group did. In the step-through avoidance task, the latency to enter the shock compartment was shorter among offspring exposed to prenatal stress than in offspring not exposed to prenatal stress. Offspring from the prenatal stress with maternal exercise group exhibited a longer latency to enter the shock compartment compared to that exhibited by their counterparts from the prenatal stress group.

Behavioral tests. (A) Spatial learning record from day 1 to day 5. (B) Morris water maze tracking. (C) Elevated plus maze tracking. (D) Time in the probe quadrant. (E) Latency in step-through avoidance task. (F) Time spent in open arms. (G) Entries into open arms. Data are presented as mean±standard error of the mean. CON, offspring born in the control group; CON+EX, offspring born in the maternal exercise group; Stress, offspring born in the prenatal stress group; Stress+EX, offspring born in the prenatal stress with maternal exercise group. *P<0.05 compared to the control group. #P<0.05 compared to the stress group.
DCX and BrdU-positive cells in the dentate gyrus
The results of DCX and BrdU-positive cells in the dentate gyrus are shown in Fig. 2. DCX and BrdU-positive cells were significantly suppressed in offspring exposed to prenatal stress in comparison to their counterparts that were not exposed to prenatal stress. Offspring belonging to from prenatal stress with maternal exercise group showed increased DCX and BrdU-positive cells compared to those exhibited by the offspring in the prenatal stress group. Offspring from the maternal exercise group did not show a significant increase in DCX and BrdU-positive cells compared to those observed in the offspring from the control group.

Number of DCX-positive and BrdU-positive cells in the dentate gyrus. (A) Photomicrograph of DCX-positive cells. (B) Photomicrograph of BrdU-positive cells. (C) Number of DCX-positive cells in each group. (D) Number of BrdU-positive cells in each group. The scale bar represents 500 μm in each group. Data are presented as mean±standard error of the mean. DCX, doublecortin; BrdU, 5-bromo-2’-deoxyuridine; CON, offspring born in the control group; CON+EX, offspring born in the maternal exercise group; Stress, offspring born in the prenatal stress group; Stress+EX, offspring born in the prenatal stress with maternal exercise group. *P<0.05 compared to the control group. #P<0.05 compared to the stress group.
BDNF, PSD-95, and synaptophysin expression in the hippocampus
The expression levels of BDNF, PSD-95, and synaptophysin in the hippocampus are shown in Fig. 3. BDNF, PSD-95, and synaptophysin expression levels were significantly suppressed in offspring exposed to prenatal stress compared to the expression levels observed in their counterparts not exposed to prenatal stress. The offspring in the prenatal stress with maternal exercise group showed increased BDNF, PSD-95, and synaptophysin levels compared to those in the prenatal stress group.
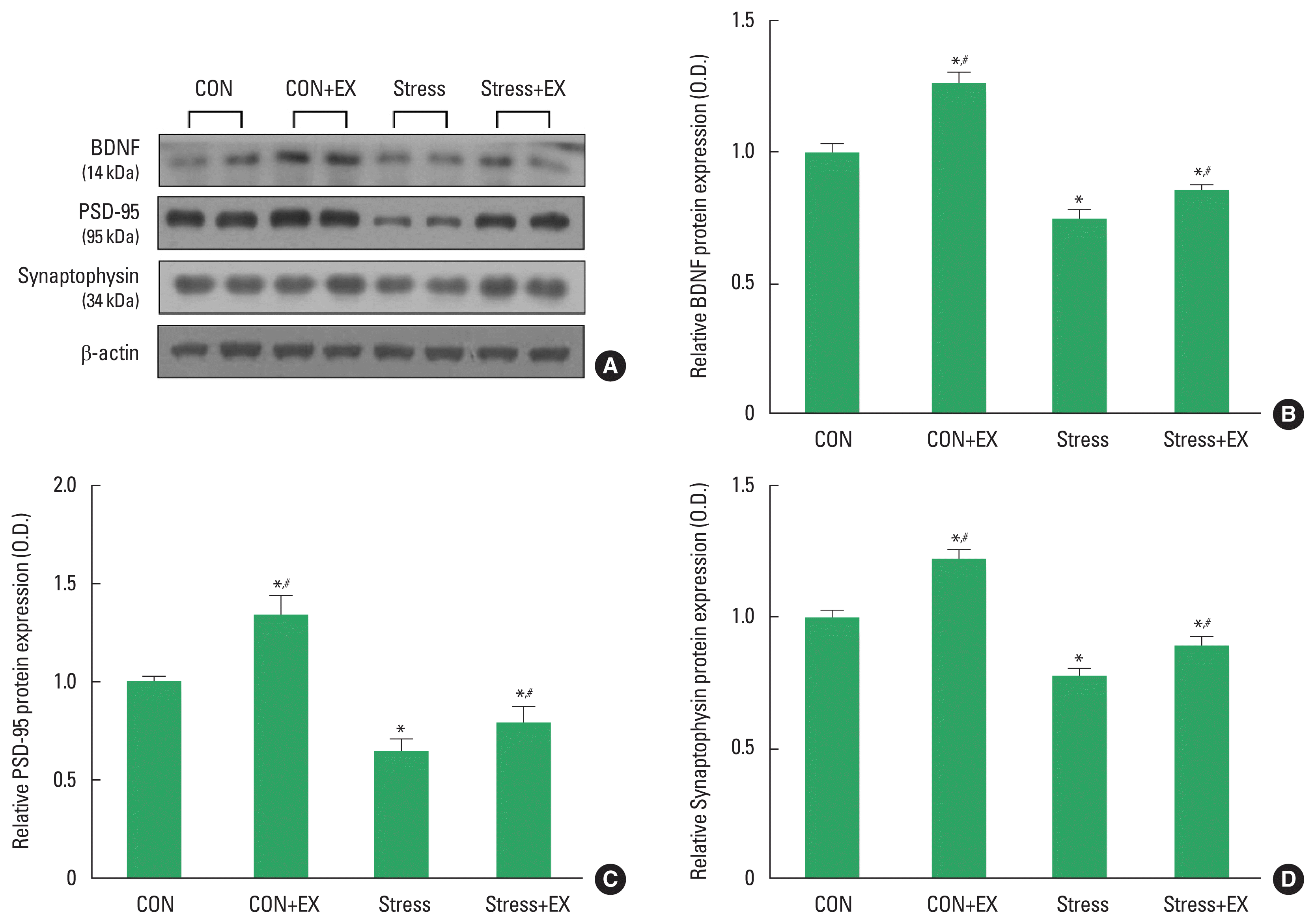
Expressions of BDNF, PSD-95, and synaptophysin in the hippocampus. (A) Representative expression of BDNF, PSD-95 and synaptophysin. (B) Density of BDNF expression in each group. (C) Density of PSD-95 expression in each group. (D) Density of synaptophysin expression in each group. Data are presented as mean±standard error of the mean. BDNF, brain-derived neurotrophic factor; PSD-95, postsynaptic density 95 kDa; CON, offspring born in the control group; CON+EX, offspring born in the maternal exercise group; Stress, offspring born in the prenatal stress group; Stress+EX, offspring born in the prenatal stress with maternal exercise group. *P<0.05 compared to the control group. #P<0.05 compared to the stress group.
Mitochondrial oxygen respiration in the hippocampus
The results of mitochondrial oxygen respiration are shown in Fig. 4. Mitochondrial oxygen respiration in complex I substrate was reduced in offspring exposed to prenatal stress compared to their counterparts that were not exposed to prenatal stress. Offspring from the prenatal stress with maternal exercise group showed an increase in oxygen consumption compared to offspring from the prenatal stress group. Mitochondrial oxygen respiration in complex I substrate and state 3 conditions did not differ significantly among the groups. Mitochondrial oxygen respiration in complex I, state 3 condition, and complex II substrate was significantly decreased in offspring exposed to prenatal stress compared to those observed in offspring not exposed to prenatal stress. Offspring from the prenatal stress with maternal exercise group showed an increase in oxygen consumption compared to that observed in their counterparts from the prenatal stress group.
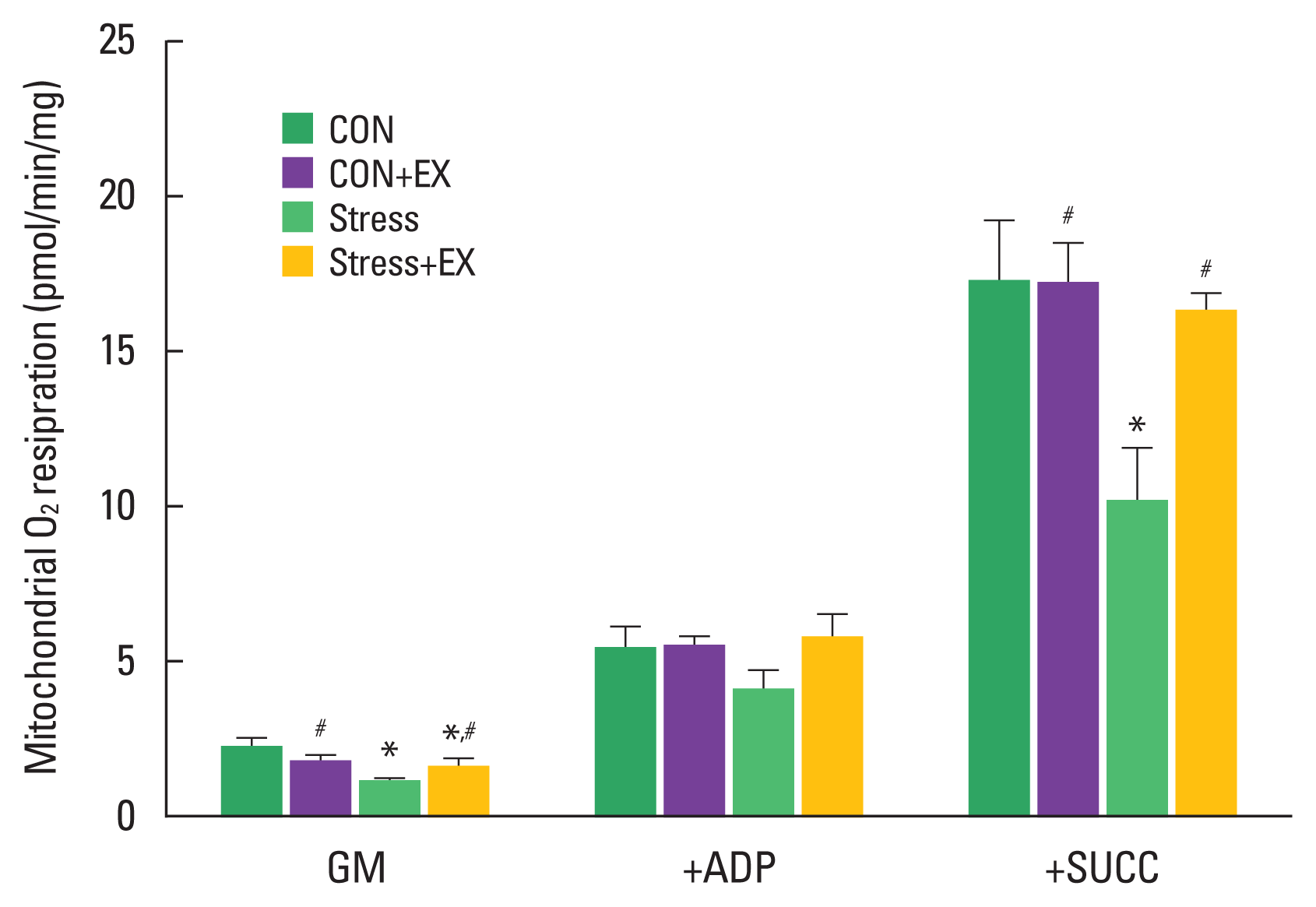
Mitochondrial oxygen consumption. Data are presented as means± standard errors of the mean. CON, offspring born in the control group; CON+EX, offspring born in the maternal exercise group; Stress, offspring born in the prenatal stress group; Stress+EX, offspring born in the prenatal stress with maternal exercise group. GM, glutamate and malate; ADP, adenosine diphosphate; SUCC, succinate. *P<0.05 compared to the control group. #P<0.05 compared to the stress group.
DISCUSSION
Prenatal stress results in learning and memory deficits associated with inhibition of hippocampal neurogenesis (Lemaire et al., 2000). Prenatally stressed rats exhibit impaired working memory and spatial learning memory during the Morris water maze test (Bustamante et al., 2020; D’Hooge and De Deyn, 2001). In this study, prenatal stress had a negative effect on spatial learning memory and short-term memory compared to fetuses not exposed to prenatal stress. Stress can negatively affect mental health, leading to anxiety and depression (Vyas et al., 2004). Głombik et al. (2015) reported that offspring exposed to prenatal stress showed fewer entries into the open arms and more entries into the closed arms, suggesting that anxiety-like symptoms were enhanced by prenatal stress. The elevated plus maze test is widely used to evaluate anxiety-like behaviors resulting from prenatal stress in rodents (Weinstock, 2017). Treadmill exercise before ischemic attack alleviated hypoperfusion-induced impairment of short-term memory in rats (Lee et al., 2019). Klein et al. (2019) reported that offspring born to exercised rats prevented amyloid-β-induced cognitive impairment. Farzad et al. (2020) reported that offspring born to exercised rats exhibited enhanced short-term memory. In our study, maternal exercise before pregnancy prevented prenatal stress-induced spatial and short-term memory deficits in offspring.
BDNF and synaptic plasticity-related proteins are closely associated with stress responses (Vollmayr et al., 2001). BDNF is a neurotrophic factor that is abundant in the hippocampus and has well-known effects on synaptic plasticity and long-term potentiation (Egan et al., 2003). Additionally, PSD-95 and synaptophysin play important roles in the regulation of synaptic function and plasticity (Rapp et al., 2004). Prenatal stress is known to inhibit BDNF, PSD-95, and synaptophysin. Moreover, it alters hippocampal synaptic plasticity and inhibits neurogenesis (Lucassen et al., 2006). In this study, prenatal stress significantly reduced hippocampal BDNF, PSD-95, and synaptophysin level in offspring exposed to stress during pregnancy compared with offspring not exposed to prenatal stress. Conversely, the offspring of mice stressed during pregnancy showed significant increase in hippocampal BDNF, synaptophysin, and synaptophysin expression by maternal exercise before pregnancy.
Reduced hippocampal cell proliferation and DCX expression are associated with prenatal stress (Lee et al., 2016). Reduced hippocampal neurogenesis is closely associated with neuropsychological disorders, and prenatal stress reduces cell proliferation and suppresses DCX expression (Bustamante et al., 2020). In our study, the number of BrdU-positive and DCX-positive cells was decreased by prenatal stress, indicating that prenatal stress inhibits new cell generation. Several studies have reported a relationship between maternal exercise and offspring hippocampal neurogenesis (Kim et al., 2007; Lee et al., 2006). Lee et al. (2006) reported that maternal exercise can alter hippocampal neurogenesis in offspring. Likewise, our results showed that exercise before pregnancy significantly increased the number of BrdU-positive and DCX-positive cells in prenatally stressed mice. Thus, exercise before pregnancy increased new cell production in offspring exposed to prenatal stress.
Hollis et al. (2015) reported that decreased mitochondrial respiratory capacity and ATP production in the hippocampus of a rat model increased anxiety. Reduced mitochondrial oxygen respiration is associated with reduced mitochondrial function, including reduced ATP production (Heo et al., 2017). In our study, offspring exposed to prenatal stress had decreased mitochondrial oxygen uptake, indicating reduced mitochondrial function and ATP production. Exercise enhances the co-expression of various enzymes involved in aerobic energy production in mitochondria (Marques-Aleixo et al., 2015). Exercise improves the expression of proteins related to mitochondrial function and reduces the expression of proteins related to oxidative stress (Marosi et al., 2012). Moreover, exercise has antioxidant properties and protects neurons from oxidative stress caused by reactive oxygen species (Lawlor and Hopker, 2001). In our study, exercise before pregnancy increased mitochondrial oxygen consumption in offspring exposed to prenatal stress. Therefore, exercise before pregnancy may alleviate decreased mitochondrial oxygen consumption in offspring exposed to prenatal stress.
Exercise before pregnancy alleviated prenatal stress-induced impairment of memory, neurogenesis and mitochondrial function. Therefore, exercise before pregnancy may have a protective effect against prenatal stress of the offspring.
ACKNOWLEDGMENTS
This work was supported by the Ministry of Education of the Republic of Korea and the National Research Foundation of Korea (NRF-2020S1A5B5A16084194).
Notes
CONFLICT OF INTEREST
No potential conflict of interest relevant to this article was reported.