INTRODUCTION
Doxorubicin (DOX) is a widely used chemotherapy drug for a variety of cancers and has demonstrated efficacy in solid tumors. However, DOX can cause cognitive decline and cardiovascular toxicity due to toxicity on brain, heart, and muscles. Cancer chemotherapy is known to cause cognitive decline and depression (Vitali et al., 2017). Chemobrain refers to a condition that causes thinking and memory problems during or after chemotherapy (Vardy and Tannock, 2007).
The hippocampus, a complex brain structure, connects with other brain regions and plays an important role in memory consolidating and memory maintenance (Nadel and Peterson, 2013). Chemotherapy causes cognitive impairment by reducing hippocampal volume and inhibiting nerve conduction (Brown et al., 2021; Christie et al., 2012). DOX itself does not cause direct damage to the brain, but indirectly causes brain toxicity (Andryszak et al., 2018). According to Yang et al. (2015), DOX forms tumor necrosis factor (TNF)-α in the brain, generates reactive oxygen species and anion radicals. Increased inflammatory substances and neurotoxicity in the brain are considered major factors contributing to decreased cognitive function and increased depression-related symptoms. Oxidative damage increases TNF-α production, which contributes to mitochondrial dysfunction and neuronal cell death (Keeney et al., 2015).
Wnt/β-catenin signaling is involved in learning and memory by regulating synaptic function and neuroplasticity (Rosso and Inestrosa, 2013). Wnt/β-catenin signaling is known to affect cognitive function by regulating neurotransmitters by acting on synaptic damage, neurogenesis, and apoptosis. DOX injection caused mitochondrial dysfunction and promotes apoptosis along with an increase of oxidative stress (Kitamura et al., 2015). Neuroplasticity is especially influenced by brain-derived neurotrophic factor (BNDF). Decreased BNDF level is associated with memory impairment. BDNF and its receptor, tropomyosin receptor kinase B (TrkB), are known to play a role in regulating cognitive function and depression (Gómez-Pinilla et al., 2002).
Regular exercise increases BDNF and synaptogenesis and improves neuronal function (van Praag et al., 1999). Exercise increases cerebral metabolism and nerve cell activation and restores cognitive function (Parker et al., 2017). The aim of this study was to investigate the impact of exercise on chemotherapy-induced memory impairment. We assessed whether DOX affects inflammation, mitochondrial Ca2+ retention capacity, and Wnt/β-catenin signaling.
MATERIALS AND METHODS
Experiment animals
Male Sprague-Dawley rats, 8 weeks old and 200 g weighing, were used in this experiment. Experimental procedures followed the regulations of the National Institutes of Health and the guidelines of the Korean Academy of Medical Sciences. This study was approved by the Institutional Animal Care and Use Committee of Kyung Hee University (Seoul, Korea) (KHUASP [SE]-20-634). Rats were randomly divided into four groups (n=10 in each group): control group, exercise group, DOX-injection group, and DOX-injection and exercise group.
DOX-induced memory impairment
We obtained DOX from Tokyo Chemical Industry (Tokyo, Japan). As a model of DOX-induced memory impairment, DOX (2 mg/kg) dissolved in saline was injected intraperitoneally once a week for 4 weeks as previously described (Park et al., 2019).
Exercise protocol
As described previously (Park et al., 2019), after a week, rats in the treadmill exercise groups were allowed to run on the treadmill for 30 min once a day, 5 days a week, for 8 consecutive weeks. The exercise load consisted of a speed of 3 m/min for the first 5 min and a speed of 8 m/min for the last 25 min.
Step-down avoidance test
A step-down avoidance test was performed to evaluate short-term memory, as described previously (Shin et al., 2019). Animals were placed on a 7×25-cm platform with a height of 2.5 cm and left on the platform for 3 min. The platform faced a 42×25-cm grid consisting of parallel steel bars of 0.1 cm aperture spaced 1 cm apart. In training sessions, rats received a 1 mA scrambled foot shock for 2 sec immediately after dismounting. After 2 hr of training, the latency (sec) of each group was measured. Latency exceeding 300 sec was counted as 300 sec.
Concentrations of proinflammatory cytokines
Concentrations of proinflammatory cytokines, such as TNF-α and IL-6, were detected by enzyme-linked immunoassay using an enzyme immunoassay kit (Abcam, Cambridge, UK) as previously described (Kim et al., 2021).
Western blotting
Hippocampal tissue was homogenized on ice using a homogenizer and lysed in lysis buffer, as previously described (Shin et al., 2019). Protein content was measured using the Bio-Rad colorimetric protein assay kit (Bio-Rad, Hercules, CA, USA). Protein was separated on an SDS-polyacrylamide gel, transferred to a nitrocellulose membrane, and incubated with mouse β-actin, BDNF, TrkB, Wnt, and β-catenin antibody. Horseradish peroxidase-conjugated anti-mouse for β-actin, BDNF, TrkB, Wnt, and β-catenin was used as the secondary antibody.
Mitochondria Ca2+ retention capacity
To measure the patency of the permeability transition pore (PTP), mitochondrial Ca2+ retention capacity was determined following a previously described method (Park et al., 2019). Mitochondrial Ca2+ retention capacity was assessed using a Spex Fluormax 4 spectrofluorometer. We measured overlapping traces of Ca2+-induced fluorescence changes by calcium green-5N by continuously monitoring ΔF/min at 37°C during state 4 conditions. After determining background ΔF, the reaction was initiated by adding Ca2+ pulses (30 μM) with innervation and emission wavelength at 506 and 532 nm. Mitochondrial Ca2+ retention capacity prior to PTP opening (Ca2+ release) was expressed as picomole/mg tissue weight.
Data analysis
To confirm the expressions of BDNF, TrkB, Wnt, and β-catenin, the detected bands were counted by densitometry. IBM SPSS Statistics ver. 26.0 (IBM Co., Armonk, NY, USA) was used for statistical analysis. Data were evaluated using one-way analysis of variance followed by Tukey post hoc test. All values are expressed as mean±standard error of the mean. P-value less than 0.05 was considered significant.
RESULTS
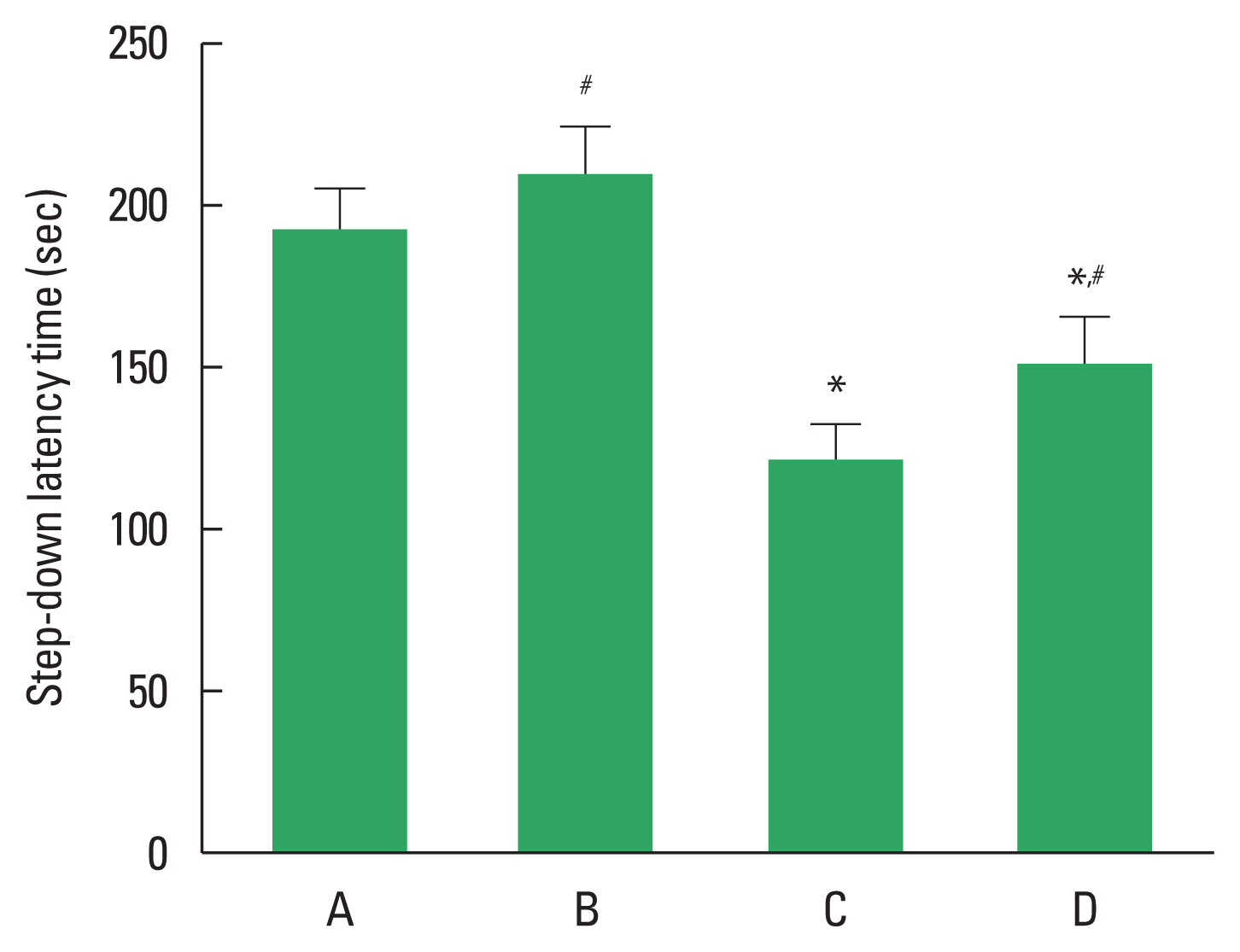
Short-term memory
Fig. 1 shows the results of step-down avoidance test. Short-term memory was deteriorated in the DOX-injected rats (P<0.05). However, treadmill exercise alleviated short-term memory impairment in the DOX-injected rats (P<0.05).
TNF-α and IL-6 concentrations
Fig. 2 shows the concentrations of TNF-α and IL-6 in in the hippocampus. Concentrations of TNF-α and IL-6 were enhanced in the DOX-injected rats (P<0.05). However, treadmill exercise suppressed concentrations of TNF-α and IL-5 in the DOX-injected rats (P<0.05).
Wnt and β-catenin expressions
Expressions of β-catenin and Wnt in the hippocampus were analyzed (Fig. 3). Expressions of phosphorylated β-catenin and Wnt were increased in the DOX-injected rats (P<0.05). However, treadmill exercise decreased phosphorylated β-catenin and Wnt expressions in the DOX-injected rats (P<0.05).
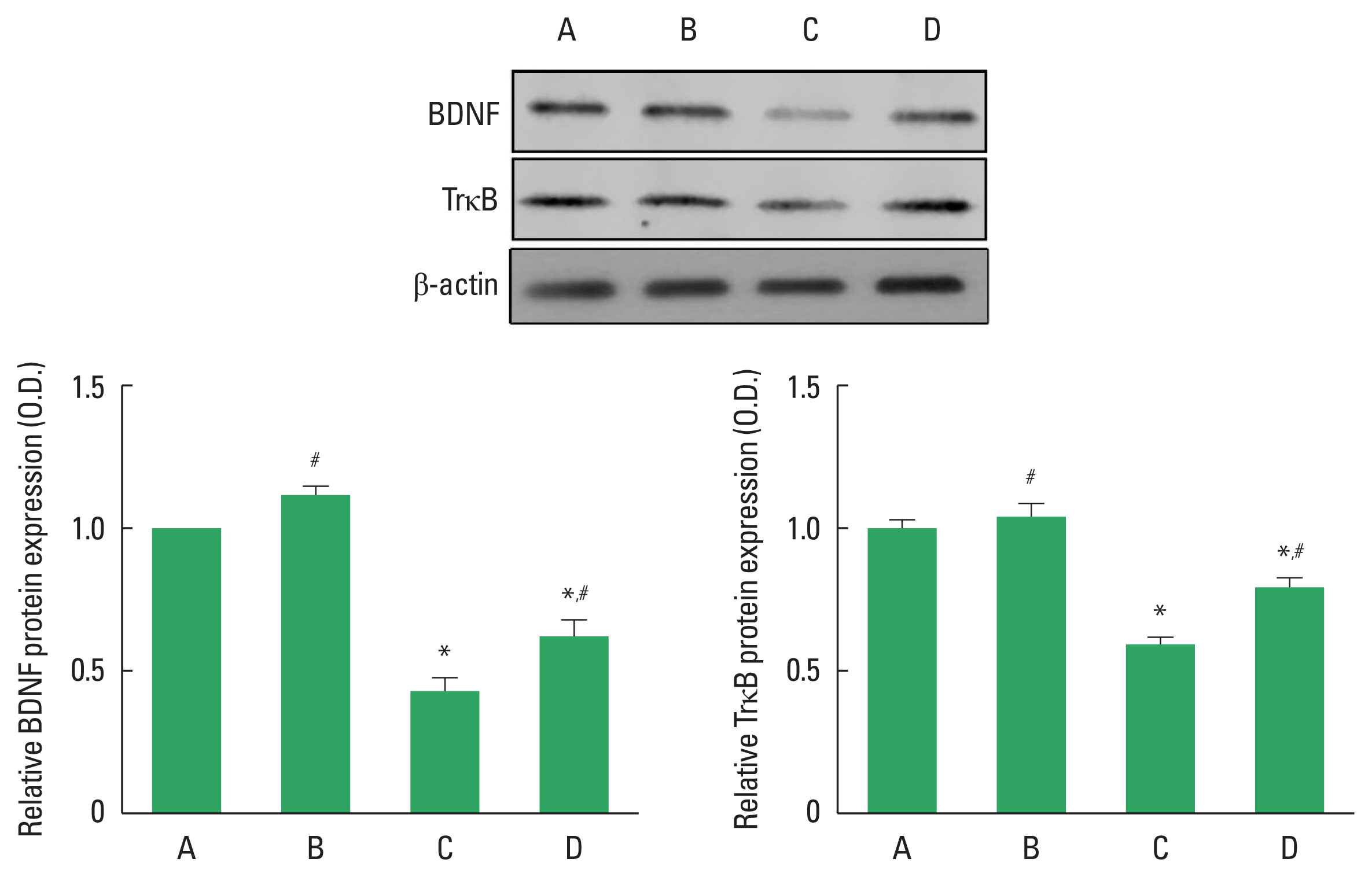
BDNF and TrkB expressions
Expressions of BDNF and TrkB in the hippocampus were analyzed (Fig. 4). Expressions of BDNF and TrkB were decreased in the DOX-injected rats (P<0.05). However, treadmill exercise increased expressions of BDNF and TrkB in the DOX-injected rats (P<0.05).
Mitochondrial Ca2+ retention capactity
Fig. 5 confirms the results of mitochondrial Ca2+ retention capacity in the hippocampus. Mitochondrial Ca2+ retention capacity was decreased in the DOX-injected rats (P<0.05). However, treadmill exercise increased mitochondrial Ca2+ retention capacity in the DOX-injected rats (P<0.05).
DISCUSSION
DOX causes short-term memory decline and cognitive impairment (Eide and Feng, 2020). In this study, short-term memory was deteriorated in the DOX-administrated rats, similar as previous study (Alharbi et al., 2020). Treadmill exercise restored reduced short-term memory in the DOX-administrated rats.
DOX causes neuroinflammation, oxidative stress, endoplasmic reticulum stress, and behavioral changes (Liao et al., 2018). DOX injection increased TNF-α and IL-6 expressions, resulting in cognitive decline (El-Agamy et al., 2018; Ren et al., 2017). Meanwhile, aerobic exercise reduced concentrations of inflammatory cytokines (Dvořáková-Lorenzová et al., 2006). Exercise suppressed hippocampal cytokine levels and enhanced antioxidant defense mechanism (Jahangiri et al., 2019). In this study, DOX injection enhanced TNF-α and IL-6 expressions in the hippocampus, in contrast, treadmill exercise suppressed TNF-α and IL-6 expressions in the DOX-injected rats.
The Wnt/β-catenin signaling pathway is a key mechanism and essential pathway for maintaining homeostasis through cell proliferation, development, migration, apoptosis, and genetic stability (Kahn, 2014; Nusse and Clevers, 2017). Wnt/β-catenin signaling acts on synaptic damage, neurogenesis, and apoptosis (Kahn, 2014; Zhang et al., 2011). In this study, DOX injection increased the expressions of Wnt and phosphorylated β-catenin, which is an overactivation of Wnt/β-catenin signaling. Meanwhile, treadmill exercise decreased expressions of Wnt and phosphorylated β-catenin, indicating restoration of Wnt/β-catenin signaling.
BDNF improves cognitive function in patients receiving anticancer drugs (Colucci-D’Amato et al., 2020). BDNF is an effective regulator of the nervous, endocrine, and immune system and is associated with nerve repair and survival, dendritic and axonal growth, and organ strengthening. Moreover, a positive correlation has been reported between BDNF level and cognitive function improvement in cancer patients (Christie et al., 2012). BDNF and TrkB expressions were decreased in DOX-injected rats, but BDNF and TrkB expressions were increased during treadmill exercise in DOX-injected rats.
DOX affects cellular oxidative stress, specifically causing mitochondrial dysfunction and neuronal mitochondrial toxicity (Andryszak et al., 2018; Yang et al., 2015). In this study, the DOX-injected rats showed mitochondrial calcium dysfunction, contributing to reduced mitochondrial Ca2+ retention capacity. In contrast, treadmill exercise improved mitochondrial Ca2+ retention capacity in DOX-injected rats. This suggests that DOX is a major cause of memory impairment through calcium dysregulation.
This study demonstrated that DOX-induced memory impairment was caused by brain inflammation and mitochondrial dysfunction through overactivation of Wnt/β-catenin signaling. Treadmill exercise ameliorated memory impairment by suppression of inflammation and restoration of mitochondrial function through normalization of Wnt/β-catenin signaling. Therefore, the present study demonstrated treadmill exercise can be used to ameliorate memory impairment in patients undergoing chemotherapy with DOX.